
Титульный лист и исполнители
РЕФЕРАТ
Отчёт 75 страниц, 30 таблиц, 7 рисунков, список использованной литературы – 143 источника.
ПШЕНИЦА, МНОГОЛЕТНЯЯ, ЯРОВАЯ ФИОЛЕТОВОЗЕРНАЯ, ПЫРЕЙ СИЗЫЙ, СОРТ, СЕМЕНА, КАЧЕСТВО ЗЕРНА, АНТИОКСИДАНТНЫЕ СВОЙСТВА, КОРМОВАЯ ЦЕННОСТЬ, ФУНКЦИОНАЛЬНОЕ ПИТАНИЕ, ЭКОЛОГИЯ, ОРГАНИЧЕСКОЕ ЗЕМЛЕДЕЛИЕ.
Объектом исследования являются сорта, гибриды, популяции и линии многолетней и фиолетовозерной мягкой пшеницы.
Цель работы – создать исходный материал для выведения сортов многолетней и яровой фиолетовозерной пшеницы, расширяющих возможности формирования внутреннего и экспортного потенциала российского рынка экологически чистого зерна, пригодного для создания безопасных и качественных, в том числе функциональных продуктов питания, корма для птицеводства и КРС.
Методы исследования: полевой и лабораторный.
Полученные результаты.
1. Проведена комплексная оценка линий многолетней пшеницы, полученными скрещиванием мягкой пшеницы с различными видами пырея и выделены лучшие для дальнейшей селекции.
2. Сорт пырея сизого Сова (альтернатива многолетней пшеницы) передан в ГСИ для включения в Госреестр, полученные оригинальные семена высеяны в семеноводческих хозяйствах степной зоны и на опытном поле Омского ГАУ.
3. Выделены лучшие клоны доместицированного пырея сизого с массой 1000 зерен не менее 10-12 г., с содержанием белка не менее 18 % для формирования синтетических популяций.
4. Проведена оценка зерна пырея сизого сорта Сова по содержанию белка, клейковины и питательной ценности для здорового питания и корма для птицеводства.
5. Разработана технология возделывания сорта Сова.
6. Проведено экологическое испытание сорта Сова в степной зоне Омской области (на базе КФХ «Тритикум»)
7. С фиолетовой окраской зерна 79 линий, испытаны в селекционном питомнике второго года по урожайности, устойчивости к болезням, оценены по качеству зерна, содержанию антиоксидантов в зерне, питательной ценности для птицеводства. Лучшие отобранные линии будут переданы в контрольный питомник.
1 Введение
Опустынивание и деградация земель являются серьезными проблемами в мировом земледелии. В 2015 году, согласно оценкам, 25 % пахотных земель в мире быстро деградируют (Nkonya et al., 2016). Основными источниками урожая в мировом земледелии являются такие культуры, как пшеница (Triticum aestivumL.), рис (Oryza sativaL.) и кукуруза (Zea maysL.). Производство однолетних монокультурных культур оказывает негативное воздействие на окружающую среду, включая загрязнение воды, эрозию почвы, сокращение накопления углерода, увеличение выбросов парниковых газов и внесение большого количества удобрений (Monfreda et al., 2008). Однолетние культуры более уязвимы, чем многолетние культуры, к эрозии почвы из-за отсутствия непрерывного почвенного покрова (Gantzer et al., 1990). Потери азота из-за однолетних культур могут быть в 30-50 раз выше, чем потери, вызванные многолетними культурами (Randall et al., 2001). Развитие многолетних культур, которые могут существовать на полях в течение многих лет, является одним из подходов, который был использован учеными для улучшения продовольственной безопасности.
Многолетние растения характеризуются способностью отрастать после урожая. Такие растения обеспечивают больший почвенный покров и имеют более продолжительный вегетационный период, чем однолетние культуры; они также обладают обширной корневой системой в почве. Экологические преимущества многолетних культур включают уменьшение эрозии почвы, защиту водных ресурсов, минимизацию вымывания питательных веществ, повышенное удержание углерода в почве и обеспечение постоянного обитания диких животных (Cox et al., 2010; Kantar et al., 2016). Экономические преимущества многолетних культур включают в себя снижение затрат на семена и удобрения (поскольку посевы производятся один раз на несколько лет), а также снижение затрат на борьбу с сорняками, обработку почвы и другие агротехнические приемы, связанные с однолетними культурами. Многолетние культуры могут использоваться не только для производства продовольствия и питания, но также для получения топлива и других непищевых биопродуктов (Colmer et al., 2006; Sanderson et al., 2008; Borrill et al., 2014; Cooney et al., 2017). Thinopyrum spp. дикий злак привлекательный в качестве многолетних доноров, благодаря своей генетической близости с Triticum spp. имеет широкое применение в селекции (Sun., 1981; Li et al., 2008; DeHaan et al., 2014). Род Thinopyrum состоит из около 11 видов с широким спектром геномного состава от диплоидов до аутоаллодекаплоидов, например Th. elongatum (2n = 2x = 14), Th. bessarabicum (2n = 2x = 14), Th. junceiforme (2n = 4x = 28 или 2n = 6x = 42), и Th. ponticum (2n = 10x = 70). Эти виды долгое время считались важными генетическими ресурсами для улучшения пшеницы, потому что содержат множество генов устойчивости к биотическим (например, болезням и вредителям) и абиотическим (солевыносливости, засухе и экстремальным температурам) стрессам (Culman et al., 2018; Li et al., 2006; Chen 2005; Li and Wang 2009; Gazza 2016). По сравнению с другими многолетними травами, Thinopyrum spp. имеет желательные агрономические признаки, включая большую массу тысячи зерен – до 5,3 г., и высокие питательные свойства зерна (Wagoner, 1990; Becker et al., 1991; Cao et al., 2007; Murphy et al., 2009). Thinopyrum spp. производит больше биомассы, чем однолетняя пшеница, и считается самым продуктивным кормовым видом на западе США (Gelfand et al., 2013; Harmoney, 2015). Thinopyrum spp. также имеет мощную корневую систему, которая способна захватывать удобрения и значительно снижать выщелачивание нитратов (Culman et al., 2018). Качество зерна Th. intermedium было таким же, как у пшеницы, с высоким содержанием белка, а мука, которая получена из данного зерна хорошо способна заменить обычную пшеницу (Jungers et al., 2017, Newell and Hayes, 2017). Ученые доказали (Larkin et al., 2014), что злак-Th. elongatum и полученные с ним гибриды с участием обычной пшеницы смогли сохраняться в поле и производить зерно более четырех лет, однако, доходность имела тенденцию уменьшаться со временем.
Ранние попытки гибридизовать пшеницу и пырей были проведены в 1920-х и 1930-х годах, когда ученые из бывшего СССР, США, Германии и Канады провели скрещивание между пшеницей и пыреем (Cox et al., 2010; Armstrong, 1936; Peto, 1936; Smith, 1943; Цицин, 1965; Scheinost et al., 2001). Первый пшенично-пырейный гибрид был получен Цициным (Цицин, 1965), который стремился развивать многолетнюю пшеницу, однако, его попытка не удалась. Тем не менее, эти исследования продемонстрировали, что возможно напрямую внедрить гены, ее придающие многолетний образ жизни в пшеницу, путем рекомбинаций или хромосомных транслокаций. Первые попытки вырастить многолетние сорта пшеницы были безуспешны до коммерческого выпуска первого многолетнего сорта пшеницы, Монтана-2 (МТ-2), в 1987 году (Schulz-Schaeffer and Haller 1987; Jones et al., 1999). МТ-2 был разработан путем скрещивания твердой пшеницы (Triticum turgidum) и Th. intermedium в университете штата Монтана в Бозмене, Монтана, США. Ученые (Lammer et al., 2004) сообщили, что дополнительная пара хромосомы 4E от Th. elongatum в сорте яровой пшеницы Chinese Spring была связана со способностью отрастать после сбора урожая; но рост был не таким сильным, как у многолетнего амфиплоидного предшественника. Сообщалось, что способность к многолетнему росту является полигенным признаком, контролируемым множеством генов (Cox et al., 2010; Kantar et al., 2016; Bell et al., 2010; Zhao et al., 2012). Это одна из трудностей при использовании Thinopyrum spp. как вид донора для развития многолетних сортов пшеницы через межвидовую гибридизацию. Вероятно, легче перенести просто унаследованные признаки одомашнивания от пшеницы в существующие многолетние виды, чтобы можно было улучшить такие дикие признаки, как признаки вымолачиваемости семян и колосьев, равномерное созревание и крупность зерна (Pinhasi van-Oss et al., 2014). Это позволит адаптировать дикие многолетние виды к современному сельскохозяйственному производству. Значительный прогресс достигнут в прямом одомашнивании нескольких многолетних видов, в том числе Th. intermedium Институт Земли (Salina, KS, USA). Двадцать перспективных многолетних линий пшеницы, разработанные на основе скрещивания пшеницы или твердой пшеницы и Th. intermedium были выращены и оценены в девяти странах мира (Cox et al., 2010; DeHaan et al., 2014). В Австралии более 150 производных пшеницы, происходящих из коллекций пшеницы Австралии, США и Китая, были оценены на способность отрастать после сбора урожая и давать урожай зерна в течение нескольких лет. Несколько многолетних линий были способны производить зерно в течение трех последовательных лет, а некоторые линии могли производить как корм, так и зерно (Bell et al., 2008; 2010; Larkin et al., 2014; Hayes et al., 2012).
Одомашнивание диких разновидностей Triticum является одним из подходов к развитию многолетней пшеницы, и в этом процессе были идентифицированы несколько ортологичных генов, которые способствуют признакам одомашнивания и улучшению однолетних культур (Kantar et al., 2016; Curwen-Mcadams and Jones, 2017; Lenser and Theißen, 2013; DeHaan et al., 2016). Например, вес зерна контролируется одним и тем же геном GW2 как в рисе, пшенице и кукурузе (Li et al., 2010, Su et al., 2011); время цветения контролируется геном VRN1 как в пшенице, ячмене и райграсе (Asp et al., 2011); а клейковина в зерне контролируется GBSSI или Waxy генами как в пшенице, ячмене, кукурузе и сорго (Kantar et al., 2016). Отбор и введение этих генов может ускорить развитие многолетних сортов пшеницы с помощью маркер-ассистированного отбора (MAS) или генетического переноса во время длительного отбора.
Одомашнивание или развитие многолетних сортов пшеницы занимает много времени и включает в себя следующие этапы: — оценка диких относительных видов и определение тех, которые обладают наибольшим потенциалом; — отбор желаемых линий до тех пор, пока не будут зафиксированы гены или локусы для одомашнивания и агрономических признаков, — интенсивное тестирование материалов в течение нескольких лет с последующим выпуском потенциальных линий (Kantar et al., 2016; DeHaan et al., 2016). Во время этого процесса агрономически отобранные растения могут быть использованы в качестве родителей для гибридизации. Дикие сородичи часто плохо справляются с агрономическими признаками из-за генетической сложности некоторых признаков и связи желательных и нежелательных признаков, таких как позднее цветение, маленький размер семян и обмолот семян. Thinopyrum spp. и другие многолетние виды в племени Triticeae могут быть гибридизованы с коммерческими сортами пшеницы, чтобы объединить многолетние привычки роста с продуктивностью пшеницы (Fradkin et al., 2012). Методы селекции, такие как подбор родословных, обратные скрещивания и повторный отбор, могут применяться для удаления вредных признаков при сохранении многолетнего признака (Kantar et al., 2016; Newell and Hayes, 2017). При отборе в раннем поколении (F2 — F4) следует выделить такие признаки, как размер семян, высота растения, самооплодотворение и хромосомный состав. Стабильному количеству хромосом также следует уделять приоритетное внимание (Kantar et al., 2016; Curwen-Mcadams and Jones, 2017). Отбор в более поздних поколениях должен фокусироваться на таких признаках, как урожай зерна и его качество, устойчивость к болезням и отрастание после сбора урожая (Marti et al., 2015, 2016; Catani, 2016). В дополнение к нескольким поколениям отбора признаков, особое внимание следует уделить генетическим изменениям во вновь разработанных аллоплоидных линиях, чтобы обеспечить согласованность между чужеродными геномами и геномами пшеницы (Larkin and Newell, 2014). Плодородие и стабильность должны учитываться в процессе развития многолетних сортов пшеницы. Помимо соображений о плоидности и генетической совместимости, многолетние линии пшеницы могут не проявлять способности к возобновлению роста в средах, отличных от той, в которой они были отобраны, из-за различий в климате (осадки и температура), почве и патогенах, которые влияют на пригодность отдельных линий (Kantar et al., 2016; Larkin and Newell, 2014).
До настоящего времени гены, которые придают многолетний образ жизни, не были идентифицированы. Исследования показали, что некоторые атрибуты многолетних привычек развития присутствуют, когда к пшенице добавляются дополнительные хромосомы от многолетних доноров (Lammer et al., 2004; Zhang et al., 1996, 2014; Jauhar, 1990; DeHaan and Van Tassel, 2014). Потенциально многолетние линии пшеницы, которые показывают многолетний тип развития, обычно содержат группу хромосом из многолетних растений. Родителям и тем, кто выжил в течение нескольких лет в полевых условиях, требовался как минимум один геном из Th. intermedium (Cox et al., 2002; Hayes et al., 2012). С увеличением хромосом пырея и уменьшением хромосом пшеницы гибриды могут улучшать признаки многолетнего образа жизни; тем не менее, нет никаких данных относительно того, какой процент хромосом из видов пырея обеспечит оптимальный многолетний образ жизни. Линии потомства с меньшим количеством пшеничных хромосом часто демонстрируют серьезную генетическую нестабильность и больше похожи на дикий злак по типу роста и развития. Оценка эффектов полных геномов с использованием продвинутых поколений невозможна из-за удаления хромосом. Ученые из института земли Канзас США пытаются понять, какие промежуточные геномы улучшат производительность многолетнего типа развития, разработав серию полных амфиплоидов с различными геномными конституциями путем скрещивания диплоидных, тетраплоидных и гексаплоидных видов Triticum с Thinopyrum spp. (Kantar et al. 2016; Curwen-Mcadams and Jones, 2017; DeHaan et al., 2014; Hayes et al., 2012). Чужеродные хромосомы в частичном амфиплоиде обычно состоят из хромосом от разных геномов (Chen 2005). Эта смесь хромосом или синтетический геном в парциальных амфиплоидах может вызывать плохую фертильность и потерю многолетних донорских хромосом. Хромосомы из разных геномов Thinopyrum spp. могут быть определены с помощью анализа GISH с использованием геномной ДНК из Pseudoroegneria provifolia или Pseudoroegneria strigosa основаная на различных моделях полос (Chen, 2005; Jones et al., 1999). Например, многолетний сорт пшеницы МТ-2 состоит из геномов A, B, D, E и St, и вариация в составе хромосом была обнаружена в пределах и внутри линий МТ-2 Чен и др. (Chen et al., 2003) сообщили, что геном MT-2 включает 10 хромосом из St генома, восемь из Js и 13 из J генома. Некоторые исследователи рекомендовали использовать диплоидные виды пырея (например, Th. Elongatum) в качестве многолетнего донора, в результате чего получился амфиплоидный гибрид с AABBEE (аналог гексаплоидного тритикале, AABBRR) или AABBDDEE (аналог октоплоидного тритикале, AABBDDRR) при использовании AABBDDRR) тетраплоидная пшеница или гексаплоидная пшеница в качестве родителей соответственно (Kantar et al.. 2016; Larkin et al., 2014; Hayes et al., 2012). Восстановление полной фертильности, повышение урожайности и способности к отрастанию может потребовать нескольких поколений селекции. Определение роли цитоплазмы в развитии многолетних растений путем межвидовой гибридизации необходимо, потому что многие исследования показали взаимодействие между ядром и цитоплазмой во время гибридизации (DeHaan and Van Tassel, 2014; Ma and Gustafson, 2008; Sykes at al., 2017). Так как дикий злак несовместим с цитоплазмой пшеницы, ген (или гены) придающие многолетний образ жизни от Thinopyrum spp. могут быть устранены или подавляться.
Быстрое развитие методов секвенирования следующего поколения (NGS) позволяет производить высококачественные эталонные генные последовательности для многих сельскохозяйственных культур и видов растений, которые могут предоставить полезную информацию для ускорения размножения многолетних злаков. С помощью NGS можно справиться с большими и сложными геномами (как диплоидными, так и полиплоидными). промежуточный тип — это аллогексаплоидный вид (2n = 6x = 42, StStJsJsJJ) с размером генома 12,6 Гб, большинство из которых (около 80–90 %) содержат повторяющиеся последовательности (Kantar et al.. 2016). Ученые (Zhang et al., 2016) оптимизировали технологию генотипирования по секвенированию (GBS) для Th. intermedium и идентифицировало много геномных маркеров в геноме Th. intermedium без эталонной геномной последовательности. Kantarski и др. (Kantarskiet et al., 2017) разработали первую интегрированную генетическую карту Th. intermedium с 21 группой сцепления, включая 10 029 маркеров GBS и охватывая 5061 сантиморган (сМ), используя семь популяций. Эта консенсусная карта показала высокую коллинеарность с ячменным геномом и была бы полезна для лучшего понимания генетического контроля многолетнего развития в Th. intermedium посредством повышения эффективности геномной селекции на улучшение признаков доместикации Th. intermedium. Высокопроизводительные платформы фенотипирования, основанные на полевых характеристиках, предоставляют точные фенотипические данные для анализа генетического контроля многолетних признаков, а также агрономических признаков, что повышает эффективность доместикации и развития многолетних культур (Kantar et al.. 2016; Araus and. Cairns, 2014).
Доступность многолетней пшеницы с мощным отрастанием, морозостойкостью и устойчивостью к засухе, которая отвечает потребностям фермеров, ограничена. Многолетнюю пшеницу следует сначала выращивать на окраинных полях в краях сельскохозяйственных угодий, прежде чем она станет более продуктивной основной культурой. Цель конечного использования многолетних сортов пшеницы должна рассматриваться как цель размножения при разработке и улучшении этой новой культуры (Weik et al., 2004). Дополнительные проблемы, связанные с многолетними культурами, включают в себя их способность превращаться в серьезные сорняки и возможность того, что они могут служить «зеленым мостом» для определенных патогенных микроорганизмов, увеличивая тем самым риск возникновения эпидемических заболеваний.
Прогресс современных геномных подходов, используемых в обычной пшенице, а также в других хорошо адаптированных однолетних культурах, будет способствовать развитию многолетней пшеницы. Интегративные методы, сочетающие в себе геномные маркеры, мощные статистические инструменты и платформы для фенотипической оценки, произвели революцию в выращивании культурных растений и одомашнивании многолетних трав. Ввиду сложного наследования и трудоемкого отбора для многолетнего роста, высокопроизводительное генотипирование, основанное на геномных подходах и методах фенотипирования, повысит эффективность точности отбора (Jia et al., 2017; Lou et al., 2017; Wang et al., 2017; Xu and Crouch, 2008). Основанные на геномике методы быстрого размножения, которые позволяют многократно производить сельскохозяйственные культуры в год (Watson et al., 2018) сократят время цикла размножения и ускорит улучшение существующих многолетних линий и одомашнивание многолетних злаковых трав.
Экологические преимущества многолетней пшеницы по сравнению с однолетними зерновыми культурами широко признаны, включают в себя: снижение эрозии почв, снижение выщелачивания нитратов почвы, увеличение поглощения углерода и сокращение затрат на семена, обработку почвы, топливо и пестициды (Glover et al., 2010; Culman et al., 2013; DeHaan et al., 2014). Многолетняя пшеница обладает устойчивостью к различным заболеваниям обычной пшеницы (Zhang et al., 2014). По содержанию лизина многолетняя пшеница не превосходит обычную, но имеет больший процент других незаменимых аминокислот. Сделанные из многолетней пшеницы кексы и печенья обладают высокими вкусовыми свойствами (Zhang et al., 2014). При добавлении в тесто 15 % муки многолетней пшеницы удалось получить объемы хлеба, сопоставимые с хлебом полученным из сильных сортов пшеницы (Wagoner, 1990). Пять генов субъединиц глютенов были идентифицированы и охарактеризованы в Thinopyrum intermedium, разделяя консервативную структуру с аналогичными в обычной пшенице (Cao et al., 2014; Zhang et al., 2014). Thinopyrum intermedium имеет большое разнообразие белковых полимеров. Некоторые генотипы почти не содержат крупных полимеров клейковины, а отдельные имеют большое количество и по качеству хлеба не уступают мягкой пшенице (Zhang et al., 2015).
Начиная с 1920-х годов, как американские, так и российские ученые изучали несколько линий многолетней пшеницы. Разновидности пырея (Agropyron glaucium и А. elongatum) были использованы академиком Н. В. Цициным для целей отдаленной гибридизации – скрещивания с пшеницей. В результате получено большое разнообразие многолетних и зернокормовых форм пшеницы.
В начале 1980-х годов институт штата Канзас начал поиск в банках семян диких злаков и опробовал около 100 видов, собранных со всего мира, прежде чем остановиться на одном из видов, называемый Thinopyrum intermedium, впервые собранный в Турции и Афганистане (Wagoner, 1990). В 2001 г. стартовал проект по изучению неисчерпаемого потенциала многолетних диких видов. При выращивании в северных условиях, что актуально для России, средняя урожайность зерна Thynopirum intermedium возросла до 1,5 т/га (DeHaan et al., 2014; Culman et al., 2013), что аналогично урожайности яровой пшеницы в Сибирском регионе. Эти многообещающие урожаи привели к созданию первого коммерческого сорта Kernza. В последнее десятилетие потенциальная урожайность Кернзы ежегодно увеличивается на 10 процентов (DeHaan et al., 2016). Полученные нами данные испытания многолетней пшеницы в Омске показывает урожайность 1,0 т/га, при этом она стабильно возрастает. С 2011 г. многолетняя пшеница стала важным селекционным направлением в США, охватывающим несколько научных отделов, включая генетику растений и агротехнологию (DeHaan et al., 2016). Программа университета штата Вашингтон, разрабатывает свою собственную версию пшеничного многолетника под названием Salish Blue. Salish Blue живет около двух лет, и фермеры в северо-западном Вашингтоне испытывают данный сорт на своих полях (Curwen-McAdams 2017). С 2001 года потенциальный размер семян многолетней пшеницы удвоился, и, ученые надеются продлить продолжительность жизни с 5 до 10 лет. В США и других странах, таких, как Канада, Австралия, Китай, в настоящий момент активно приступили к исследованиям в области генетики и селекции многолетней пшеницы. Участники различных селекционных программ пытаются создать сложные межвидовые и межсортовые гибриды для получения этих культур. Использование генотипирования и секвенирования генома позволило получить маркеры для Thynopirum intermedium (DeHaan et al., 2016). Прогнозирование генома внедряется для ускорения одомашнивания промежуточного пырея за счет повышения эффективности отбора и сокращения цикла селекции (Zhang et al., 2016). В настоящее время основные направления селекции Thynopirum intermedium устойчивость к полеганию, легкость обмолота. Изучают прочность зерна с качествами похожими на пшеницу и выявляют генетическую изменчивость, которая позволит создавать сорта для конкретных видов продукции (хлеб, блины, пиво) и для смешивания с пшеницей и повышения её качества (DeHaan et al., 2016).
Пшеница является важной культурой для продовольственной безопасности в мире и основным продуктом питания для миллионов людей как в развитых, так в развивающихся странах. В настоящее время продовольственная безопасность подразумевает не только обеспеченность зерном, но и продуктами питания из зерна пшеницы, которые положительно влияют на здоровье человека. В контексте пищевой ценности и пользы зерна для здоровья человека недавно были расставлены акценты благодаря изучению всего разнообразия возделываемой пшеницы: различных видов, таких как однозернянка и спельта, местные сорта и стародавние сорта, современные сорта и краснозернная пшеница. Ряд научных публикаций, посвященных пшенице с голубым и фиолетовым зерном, полученной за последние два года, указывает на перспективность создания сортов пшеницы, оказывающих положительное влияние на здоровье человека. Интерес к фиолетовозерной пшенице обусловлен наличием флавоноидных пигментов антоцианов, обнаруженных в самых разных растениях, включая фрукты и овощи. Антоцианы действуют как антиоксиданты, разрушая свободные радикалы [Graisgruber et al., 2018]. Исследования Sytar et al. [2018] продемонстрировали высокую корреляцию между антиоксидантной активностью и содержанием биоактивных фитохимикатов (антоциановые компоненты цианидин и пеларгонин) в окрашенном зерне. Польза антоциана для здоровья была недавно показана Daotong Li [Daotong Li et al, 2017]. Важнейшие клеточные процессы, такие как клеточный цикл, апоптоз, аутофагия и биохимический метаболизм могут способствовать терапевтическому эффекту и лечению широкого спектра заболеваний в будущем.
Происхождение генетических ресурсов пшеницы с фиолетовым зерном было описано в обзорах Zeven [1991], а недавно Grausgruber et al. [2018]. Фиолетовая тетраплоидная пшеница была впервые отобрана в Абиссинии (ныне Эфиопия) в 1870-х годах и завезена в Европу, где она была широко распространена ботаниками с повторным интродуцированием. Признак фиолетовой окраски зерна был в конечном счете интродуцирован в мягкую пшеницу. В 1960-х и 1970-х годах фиолетовозерная пшеница была получена во всем мире на кормовые цели, для определения степени перекрестного опыления. Коммерческие сорта фиолетовозерной пшеницы были созданы в Новой Зеландии, Австралии, Канаде, Китае и ряде европейских стран. Интересно, что в Эфиопии по-прежнему возделывают фиолетовозерные сорта и их можно найти на рынках в районе реки Тигр [Kassegn, 2018].
Пигментация антоцианов в пшенице контролируется доминантными аллелями генов: Rc (колеоптиль), Pc (стебель), Plb (листовая пластинка), Pls (листовое влагалище), Pp (перикарп), Pan (пыльники) и Ra (бутоны) [Хлесткина и др., 2002; 2009; 2014; Терещенко и др., 2012; 2013]. Аллели, контролирующие сильную пигментацию, обусловливают активную транскрипцию структурных генов, играя важную роль в качестве регуляторов в биосинтезе антоцианов. Согласно Терещенко и др., [2012], темно-фиолетовый перикарп является результатом комплементарного действия доминантных генов: Pp-D1 (7D) и Pp 3 (2A). Наличие доминантных аллелей Pp-A1 и Pp3 приводит к светло-фиолетовуму цвету, и зерно останется неокрашенным в присутствии рецессивного аллеля Рp3, даже если два других аллеля (PpA1 и Pp-D1) находятся в доминантном состоянии [Gordeeva et al. 2015]. Позднее комплементарное действие генов Pp3 и Pp-1 было объяснено взаимодействием продуктов синтеза (транскрипционный фактор bHLH-MYC, кодируемый геном Pp3 и фактор R2R3-MYB, кодируемый генами Pp-1), что приводит к функциональному регуляторному комплексу MBW для биосинтеза антоцианов [Shoeva et al., 2014].
Исследования в области селекции и поиск генетических ресурсов фиолетовозерной пшеницы проводятся во всем мире. Программа селекции озимой пшеницы в Республике Чехия направлена на сочетание темноокрашенного зерна с сочетанием разных генов, важных агрономических признаков и хорошего качества хлеба [Martinek et al.; 2013]. В Китае с помощью гамма-облучения было получено пять новых мутантных форм гексаплоидной фиолетовой линии пшеницы K4191 с различным цветом зерна [Hong et al., 2018]. Общее содержание антоцианов в трех мутантных линиях было значительно выше, чем в исходной и белозерной в качестве контроля. Содержание антоцианов в зерне может быть увеличено за счет пирамидирования различных генов, ответственных за накопление антоцианов в перикарпе зерновки [Bohmdorfer et al., 2018]. Они предложили метод высокоэффективной тонкослойной хроматографии, позволяющий быстро и эффективно фенотипировать пигменты в зерне. Этот же метод был успешно использован для фенотипирования 94 генотипов пшеницы с окрашенным зерном [Kryger S .; Morlock G.E., 2018]. Стоимость фенотипирования была ниже 1 евро, а время обработки — менее 9 минут на образец. Скрещивания между генотипами, имеющими фиолетовый перикарп и голубой алейроновый слой, были использованы для увеличения содержания антоцианов, хотя большинство гибридов имели признаки родительских форм (Syed Jafar et al., 2013).
Одним из основных направлений недавних исследований является содержание антоцианов в зерне пшеницы, его состав, различия в генотипах и продуктах питания. Knivel et al., (2009) изучал адаптированные линии яровой пшеницы с голубым и фиолетовым перикарпом в течение двух лет в Саскатуне (Канада) и обнаружил, что генотипические различия по концентрации антоциана были статистически значимыми, в то время как влияние фактора Год и Взаимодействия (генотип х год) статистически недостоверны. Это облегчает селекционный процесс. В недавнем исследовании девяти генотипов фиолетовозерной пшеницы в Саскачеване (Канада) по содержанию и составу антоциана существенное влияние оказывали факторы Генотип и Взаимодействия (генотип х окружающая среда) [Abdel-Aal et al., 2016]. Накопление ассимилятов в зерне играет важную роль в содержании антоцианов [Bustos et al., 2011]. Положение зерновок в колосе влияло на содержание антоцианов, в дистальных участках колоса содержание антоцианов уменьшалось на 25-50 %. Тепловой стресс положительно влиял на содержание антоцианов в зерне пшеницы [Hosseinian F.S.2008; Li et al., 2018]. Одновременное внесение азотных и фосфорных удобрений приводит к повышению антиоксидантной активности при изучении шести сортов пшеницы, включая два фиолетовозерных [Ma D., et al., 2018]. Shoeva и др. [2017] показали, что накопление антоцианов в колеоптиле защищает побег, в то время как в зернах – корневую систему в условиях засухи (когда сеянцы выращивали в 15 % PEG). Кроме того, антоцианы в колеоптиле защищают проростки пшеницы от воздействия токсичных тяжелых металлов (при обработке 25 мкМ CdCl2) [Шоева, Хлесткина 2018] и при умеренной дозе облучения (предпосевная обработка сухих семян дозой облучения 50 Гр) [Гордеева и др., 2018].
Четыре сорта озимой пшеницы с окрашенным зерном вместе с двумя стандартами были изучены в течение двух при агротехнике, характерной для традиционного и органического земледелия [Zrckova et al., 2018]. В то время как общее содержание фенолов больше зависело от года, на содержание антоцианов больше влияли генотипы, а влияние среды и генотипов было сопоставимо с общей антиоксидантной активностью. Более высокое содержание антоцианов, антиоксидантная активность и более низкие урожаи наблюдались в системе органического земледелия и в более сухой год. Тринадцать различных генотипов пшеницы с повышенным содержанием антоцианов изучали в Австрии в течение двух лет [Eticha et al. 2011]. Генотипы и факторы окружающей среды оказывали большое влияние на агрономические и биохимические признаки. Статистически достоверная и относительно высокая отрицательная корреляция наблюдалась между массой тысячи зерен, с одной стороны, и общим содержанием фенолов и антоцианов, с другой стороны. В то же время содержание белка не влияло на биохимические признаки.
Была проведена оценка качества продуктов, полученных из фиолетовозерной пшеницы, в частности пива [Li et al., 2007], печенья [Pasqualone et al., 2015; Усенко и др., 2018], макаронных изделий [Fiocco и др., 2016), хлеба [Li and Beta, 2011; Хлесткина и др., 2017], булочек [Paznocht et al., 2019), порошка из отрубей [Abdel-aal et al., 2018], китайской лапши [Ma D et al., 2018] и бездрожжевого хлеба [Tian SQ . Чен З .; 2018]. Hrivna и соавторы [2018] изучали реологические свойства теста, содержащего 10; 15 и 20 % отрубей фиолетовозерной пшеницы. Добавление отрубей увеличивало потери воды при выпечке и удельный объем хлеба. Как правило, более высокое содержание отрубей увеличивает вязкость теста. Детальная оценка свойств муки и теста из фиолетовозерной и голубозерной пшеницы позволила установить, что сорта пшеницы с окрашенным зерном не отличались по основным показателям от коммерческих сортов пшеницы [Buresova et al .; 2019]. Хлесткина и соавт. [2017] показали, что качество выпечки хлеба и органолептические свойства хлебобулочных изделий, изготовленных из фиолетовозерной пшеницы, не уступали, или в некоторых случаях превышали соответствующие свойства продуктов, полученных из контрольных зерен NIL. Было установлено, что присутствие антоцианов в зерне увеличивает срок хранения хлебобулочных изделий и их устойчивость растений к произрастанию в стрессовых условиях [Хлесткина и др., 2017].
В большинстве исследований фиолетовозерной пшеницы использовался и сравнивался очень разнообразный материал, включая стародавние и современные сорта, сорта с различной адаптацией и даже разные виды. При использовании такого разнообразного материала трудно изучить влияние Pp-генов как таковых, так как влияние генотипического фона очень существенно. В институте Цитологии и генетики в Новосибирске (Россия) получены изогенные линии с фиолетовым зерном на основе сорта Саратовская-29 [Арбузова и др., 1998; Гордеева и соавт. 2015]. Были использованы два донора генов Pp: сорта Purple и Purple Feed. Материалом в данном исследовании послужили морфологически идентичные изогенные линии Саратовская-29. Цель исследования состояла в том, чтобы сравнить изогенные линии с красным (рекуррентным родительским) и фиолетовым зерном по урожайности и компонентам урожая, признакам качества хлеба и составу веществ, оказывающих влияние на здоровье человека, в контрастных условиях выращивания для разработки соответствующих подходов в селекции.
Фиолетовые сорта пшеницы впервые были коммерчески произведены в Новой Зеландии для придания цвета и текстуры цельнозернового хлеба (Griffin, 1987). В Европе, хлебобулочные изделия из муки грубого помола, изготовленные из пурпурной пшеницы были введены в 2006 году Backaldrin (The Kornspitz Company, Asten, Австрия) и выпускались под брендом PurPur. В Канаде, фиолетовые продукты питания из пшеницы продают под брендом InfraReady Products, Саскатун, под зарегистрированной торговой маркой AnthoGrainTM (InfraReady Products Ltd., Саскачеван, Канада). В Китае, антоцианы в пигментированных сортах пшеницы были разработаны для лапши быстрого приготовления и соевого соуса (Li., 2011). Фиолетовый цвет зерна определяют гены фиолетового перикарпа Рр, которые были переданы мягкой пшенице от Triticum turgidum L. Антоцианы цианидин 3-глюкозид, цианидин 3-рутоназид и сукцинил глюкозид наиболее представлены в фиолетовых зернах (Knievel et al. 2009). Основным направлением использования фиолетовозерных сортов пшеницы является создание продукции здорового питания.
Пшеница не только считается источником белка и углеводов, но также признана источником в снижении риска хронических заболеваний, связанных с возрастом, таких как сердечно-сосудистые заболевания, нейродегенерация, диабет II типа, ожирение и некоторые виды рака, благодаря высокому содержанию антиоксидантов. Фиолетовозерная пшеница имеет большую активность антиоксидантов, чем обычная пшеница (Li et al., 2005). Результаты указывают, что фиолетовые сорта пшеницы имеют самое высокое содержание фенолов, за ними следует краснозерная и белозерная пшеницы (Liu et al., 2010). В последние годы количества антиоксидантных фитохимикатов пшеницы вызвали значительный интерес как у исследователей, так и у производителей продуктов питания. Основным антиоксидантом, содержащимся в зерне пшеницы служит ферулевая кислота и является основным фенольным соединением. Связанные фенолы считаются уникальными антиоксидантами обладающие противовоспалительными свойствами (Adom, 2002). Эти соединения помогают снизить риск развития колоректального рака, так как могут улучшить пищеварение в желудке и тонком кишечнике, и после абсорбции они улучшают работу толстого кишечника и других тканей (Liu, 2007 ). Белая пшеничная мука, в которой отсутствует большая часть отрубей, не содержит фенольных соединений (Beta et al., 2005).
2 Метеорологические условия в год проведения опытов
Исследования проводились в саду им. Кизюрина и на малом опытном поле Омского ГАУ, расположенном на правом берегу реки Иртыш, в зоне южной лесостепи Омской области. Для характеристики погодных условий в 2018 году использованы наблюдения метеорологической станции «Омск-Степная» (таблица 1).
Таблица 1 – Метеорологические данные за 2019 г.
| Месяц | Май | Июнь | Июль | Август | Сентябрь | |||||||||||
| Декада | I | II | III | I | II | III | I | II | III | I | II | III | I | II | III | |
| Температура, °C | ||||||||||||||||
| Температура 2019 г., 0С | 13,9 | 9,6 | 13,1 | 14,2 | 15,6 | 16,6 | 18,8 | 22,2 | 20,5 | 19,5 | 19,2 | 15,3 | 11,3 | 14 | 7,1 | |
| Средне многолетняя температура | 9,9 | 12,1 | 14 | 16 | 18,3 | 18,9 | 19,8 | 19,7 | 18,8 | 17,8 | 16,9 | 14,6 | 12,7 | 10,8 | 8,1 | |
| Осадки, мм | ||||||||||||||||
| Осадки
2019 г., мм |
0,3 | 13 | 24 | 52 | 21 | 11 | 23 | 0 | 6 | 13 | 12 | 15 | 22 | 2 | 23 | |
| Средне многолетние осадки, мм | 10 | 10 | 14 | 14 | 17 | 22 | 21 | 21 | 25 | 20 | 16 | 17 | 13 | 10 | 8 | |
Первая декада мая была значительно более теплой, на 4 0С выше, чем в среднем, при этом осадков практически не было.
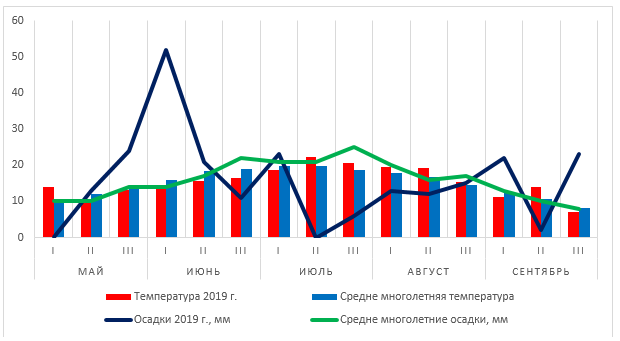
Рисунок 1 — Сравнительная характеристика осадков и температуры воздуха за 2019 г. со среднемноголетними данными
Во второй декаде мая температура воздуха была на 2,5 0С ниже нормы при превышении нормы осадков на 3 мм. Третья декада мая так же была более прохладная со значительно большим количеством осадков – 24 мм при норме 14 мм. Июнь был в целом более прохладным, с колебанием температуры по декадам от 14,2 до 16,6 0С, тогда как среднемноголетние показатели составляют 16-18,9 0С. Погодные условия июля были значительно более жесткими, лишь первая декада месяца была близка к нормальным условиям – прохладнее на 1 0С и на 2 мм с большим количеством осадков. Вторая и третья декада месяца были более жаркими – на 2,5 и 1,7 0С соответственно и практически без осадков. Август так же был более жарким, чем обычно, температура колебалась от 19,5 в первой декаде до 15,3 0С в третьей, при норме соответственно 17,8 и 14,6 0С. Осадков за месяц выпало на четверть меньше нормы. Сентябрь характеризовался прохладной и дождливой погодой в первой и третьей декадах и значительно более теплой и сухой второй декадой.
Таким образом, вегетационный период 2019 года характеризовался значительным колебанием количества осадков по декадам и близкими к нормальным значениями температуры в сравнении со среднемноголетними данными.
3 Комплексная оценка линий многолетней пшеницы, полученных скрещиванием мягкой пшеницы с различными видами пырея
Всего в коллекции многолетней пшеницы изучено 25 линий, характеристика лучших из них приведена в таблице 2.
Таблица 2 – Родословная лучших линий многолетней пшеницы коллекции Омского ГАУ
| № в коллекции | Название | Родословная |
| 2 | Agrotana | T. aestivum x Th. ponticum |
| 4 | 11955 | T. aestivum x Th. ponticum |
| 10 | 235а | T. aestivum x L. elongatum |
| 11 | TAF46 | T. aestivum x Th. intermedium |
| 12 | Ot 38 | T. aestivum x Th. intermedium |
| 14 | ОK-72 | T. aestivum x Th. ponticum |
Как видно из таблицы 2, 3 линии, выделившиеся по результатам двухлетнего изучения, созданы на основе Th. Ponticum. Две линии содержат в родословной Th. Intermedium, и одна L. Elongatum.
В таблице 3 представлены данные полевых наблюдений выделившихся образцов коллекции многолетней пшеницы.
Таблица 3 ‒ Данные полевых наблюдений выделившихся оразцов коллекции многолетней пшеницы
| № | Название | Число всходов | Перезимо-вавших | Процент перези-мовки | Выход в трубку | Колоше-ние | Мучнистая роса | Стебле-вая ржавчина | Бурая ржавчина | Септо-риоз |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| 2 | Agrotana | 22 | 15 | 70 | 18 июн. | 26 июн. | 2 | 0 | 0 | 5 |
| 4 | 11955 | 14 | 9 | 76 | 18 июн. | 25 июн. | 5 | 0 | 0 | 4 |
| 10 | 235а | 20 | 16 | 82 | 19 июн. | 25 июн. | 8 | 5MR | 0 | 7 |
| 11 | TAF46 | 23 | 16 | 69 | 19 июн. | 25 июн. | 5 | 0 | 0 | 8 |
| 12 | Ot 38 | 29 | 27 | 92 | 18 июн. | 2 июл. | 2 | 40S | 0 | 4 |
| 14 | ОK-72 | 17 | 12 | 74 | 21 июн. | 27 июн. | 3 | 0 | 0 | 5 |
В результате полевых наблюдений установлено:
-лучшей зимостойкостью обладает линия Ot 38, а худшая зимостойкость наблюдалась у образца TAF46;
— наилучшей устойчивостью к мучнистой росе обладают образцы Ot 38 и Agrotana, худшим в данном аспекте стал образец 235а
— устойчивостью к стеблевой ржавчине обладают образцы: Agrotana, 11955, TAF46 и ОK-72, у образца Ot 38 отмечено довольно сильное поражение стеблевой ржавчиной.
— при наблюдении бурой ржавчины отмечено полная устойчивость всех образцов.
— септориозом сильно поражались образцы 235а и TAF46, у остальных линий наблюдалось средняя устойчивость к септориозу.
Ниже представлены данные по основным хозяйственно ценным признакам линий многолетней пшеницы коллекции Омского ГАУ.
Таблица 4 – Основные хозяйственно ценные признаки линий многолетней пшеницы коллекции Омского ГАУ
| № | Высота растения, см | Длина верхнего междоузлия, см | Длина колоса, см | Ширина колоса, см | Число колосков | Масса колоса | Число зерен в колосе | Масса зерна с колоса, г | Масса 1000 семян | Урожай-ность, г/м2 | Кхоз. Раст. |
| 2 | 77 | 40 | 12,50 | 0,69 | 15,00 | 1,22 | 16,9 | 0,48 | 28,20 | 1,25 | 0,23 |
| 4 | 98 | 46 | 14,2 | 0,94 | 15,7 | 1,71 | 23,7 | 0,88 | 41,86 | 0,72 | 0,17 |
| 10 | 96 | 59 | 13,1 | 0,91 | 18,1 | 1,79 | 31,3 | 1,00 | 31,88 | 0,98 | 0,24 |
| 11 | 90 | 49 | 13,9 | 1,28 | 16,8 | 2,39 | 35,6 | 1,44 | 40,34 | 0,72 | 0,21 |
| 12 | 110 | 31 | 16,3 | 0,95 | 21,8 | 1,99 | 42,8 | 1,33 | 31,05 | 0,94 | 0,20 |
| 14 | 64 | 30 | 10,6 | 0,53 | 14 | 1,07 | 9,0 | 0,31 | 38,20 | 0,96 | 0,04 |
Высота растений варьировала от 64 см, у линии ОK-72 до 110 см у линии Ot 38. Длинна верхнего междоузлия была наибольшей у образцов — 235а (59 см) и TAF46 (49 см). Образцы Ot 38 и 11955 отличились наибольшей длинной колосьев – 16,3 и 14,2 см соответственно. Наибольшая масса колоса отмечена у линии TAF46 – 2,39 г, так же по этому признаку выделяется линия Ot 38 – 1,99 г. Наибольшее число зерен в колосе и масса зерна колоса так же отмечено у линий TAF46 – 35,6 шт. и 1,44 г и у Ot 38 – 42,8 шт. и 1,33 г соответственно. По массе 1000 зерен выделились линии 11955‒ 41,86 г и TAF46‒ 40,34 г Урожайность изученных образцов варьировала от 0,72 т/га до 1,25 т/га. Наибольшая урожайность отмечена у линии Agrotana.
Также был проведен анализ образцов на содержание клейковины и белка в зерне. Данные представлены в таблице 5.
Таблица 5 – Содержание белка и клейковины в зерне многолетней пшеницы
| № образца | Белок, % | Клейковина, % |
| 2 (Agrotana) | 19,22 | 37,89 |
| 4 (119550 | 20,04 | 45,0 |
| 10 (235а) | 18,55 | 39,08 |
| 11 (TAF46) | 19,69 | 43,6 |
| 12 (Ot 38) | 20,23 | 42,53 |
Как видим из таблицы все образцы показали очень высокое качество зерна.
Особенно выделяются образцы 4 (119550) и 12 (Ot 38), показавшие аномально высокие показатели качества клейковины и белка.
4 Изучение доместицированного пырея сизого
4.1 Результаты изучения сорта Сова
Опыт 1
2015 год посева. Питомник размножения с нормами и сроками посева
Посев произведен с 3 сроками и 3 нормами высева делянками площадью 10 кв. метров.
1 срок: 14 августа с междурядьем 40 см и 40 см между растениями, в трех кратной повторности
2 срок: 7 августа с междурядьем 40 см и 20 см между растениями, в трех кратной повторности
3 срок: 14 августа с междурядьем 20 см и 10 см между растениями, в трех кратной повторности
В результате проведенного опыта были получены следующие данные с 10 м2, отдельно зерно и зеленая масса в таблицах 5 и 6.
Таблица 6 – Зерновая продуктивность сорта Сова
| 2017 год посева | 1 срок
(среднее по повт) |
2 срок
(среднее по повт) |
3 срок
(среднее по повт) |
| Масса колосьев, г | 490,3 | 766,5 | 876,1 |
| Масса зерна, г | 251,4 | 407,1 | 414,6 |
| 2018 год | 1 срок
(среднее по повт) |
2 срок
(среднее по повт) |
3 срок
(среднее по повт) |
| Масса колосьев, г | 625,3 | 852,1 | 971,8 |
| Масса зерна, г | 298,7 | 440,5 | 447,6 |
| 2019 год | 1 срок
(среднее по повт) |
2 срок
(среднее по повт) |
3 срок
(среднее по повт) |
| Масса колосьев, г | 746,3 | 1001,3 | 1094,2 |
| Масса зерна, г | 348,3 | 510,7 | 519,4 |
Таблица – 7 Продуктивность зеленой массы сорта Сова
| 2017 год посева | 1 срок
(среднее по повт) |
2 срок
(среднее по повт) |
3 срок
(среднее по повт) |
| Масса зеленой массы, г | 7027 | 9014 | 9354 |
| 2018 год | 1 срок
(среднее по повт) |
2 срок
(среднее по повт) |
3 срок
(среднее по повт) |
| Масса зеленой массы, г | 9197 | 10240 | 10950 |
| 2019 год | 1 срок
(среднее по повт) |
2 срок
(среднее по повт) |
3 срок
(среднее по повт) |
| Масса зеленой массы, г | 9314 | 10425 | 11238 |
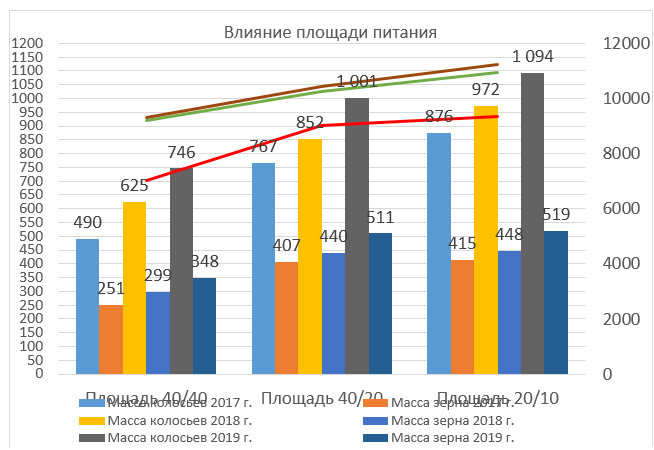
Рисунок 2 – влияние площади питания на массу колосьев и зерна.
Исходя из данных можно сделать вывод о том, что оптимальной площадью питания для растения являются 40 х 20 см, и 20 х 10 см, но экономически более эффективным является посев с площадью питания 40 х 20 см.
4.2 Выделение лучших клонов доместицированного пырея сизого для формирования синтетических популяций
Опыт 2
Работа проводилась в производственном питомнике размножения сорта Сова в 2018-19 гг.
Дисперсионный анализ отобранных высокорослых и низкорослых растений по элементам продуктивности
В питомнике размножения были отобраны 20 высокорослых и 20 низкорослых образцов, был проведен структурный анализ по одному главному колосу и осуществлен дисперсионный анализ по элементам продуктивности.
Результаты изучения представлены в таблице 8.
Таблица 8 – показатели высокорослых и низкорослых растений по элементам продуктивности.
| Высота, см | Длина колоса, см | Масса растений, г | Масса колоса, г | Масса стебля, г | Ширина колоса, мм | Число колосков колосе, шт | Масса зерна, г | Число зерен, шт. | Плотность, шт/10 см | Масса 1000 зерен, г | |
| Высоко-рослые
Среднее |
159 | 22,9 | 5,3 | 0,9 | 4,3 | 0,7 | 20,7 | 0,4 | 40,2 | 1,12 | 10,90 |
| Низко-рослые
Среднее |
107 | 19,9 | 3,0 | 0,8 | 2,3 | 0,7 | 18,2 | 0,3 | 33,8 | 1,12 | 10,23 |
| Критерий стьюден-та | 9,258E-20 | 0,004479876 | 5,27704E-07 | 0,080072152 | 5,41262E-07 | 0,151270701 | 0,01712861 | 0,120285155 | 0,258420398 | 0,905171778 | 0,611366105 |
В 2019 году были отобраны лучшие растения из 40 шт., 7 шт. низкорослых и 7 шт. высокорослых. Было посеяно с полной рандомизацией в четырехкратной повторности. Первые результаты будут получены в 2020 году. Отборы были проведены по признаку массы 1000 зерен.
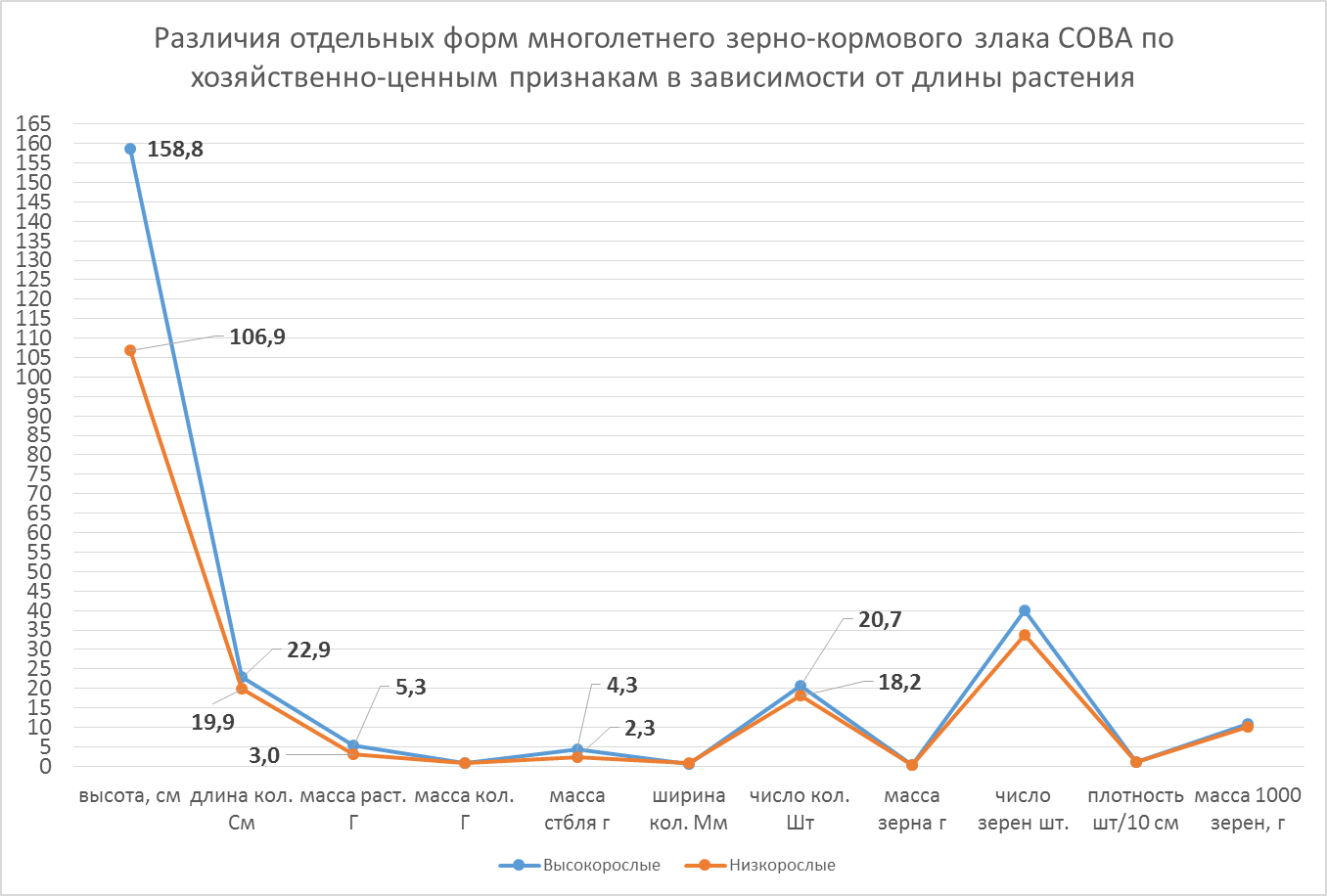
Рисунок 3 – Различия отдельных форм многолетнего зерно-кормового злака Сова по хозяйственно-ценным признакам
Исходя из данных можно сделать вывод что, положительный эффект длины растения был на признаках длины колоса, массы растения, массы стебля и числа колосков с колоса.
Высокорослые растения сорта Сова достоверно превосходили низкорослые по этим показателям.
4.3 Разработка технология возделывания сорта Сова
Опыт для установления оптимального срока укоса.
Было проведено 3 срока укоса в 4 кратной повторности с 1 квадратного метра. Данные представлены в таблице 9.
Таблица 9 – Продуктивность по срокам укоса
Таблица 9 – Продуктивность по срокам укоса
| 1 срок
(1 июля 2018г) |
2 срок
(20 июля 2018 г) |
3 срок
(1 августа 2018 г) |
|||||||
| Масса зеленых раст., г | Масса сухих раст., г | Масса зеленых раст., г | Масса сухих раст., г | Масса зеленых раст., г | Масса сухих раст., г | ||||
| Среднее по повторностям | 2312 | 537 | 3465 | 1207 | 4809 | 2038 | |||
| 1 срок
(1 июля 2019 г) |
2 срок
(20 июля 2019 г) |
3 срок
(1 августа 2019 г) |
|||||||
| Среднее по повторностям | 3252 | 743 | 5146 | 1821 | 6821 | 2846 | |||
В результате проведения опыта установлено, что оптимальным сроком укоса сорта Сова на сено является 1 декада августа.
Опыт 4
Анализ содержания антоцианина в зерне проведен на приборе Милихром-6 по методике Bicudo et al. Данные представлены в таблице 10.
Таблица 10 ‒ Анализ содержания антоцианина на приборе Милихром-6
| Тип | Пик | Содержание антоцианинов (мкг) |
| Популяция сорта Сова(фиолетовая) | 294 | 57,91 |
| Сорт Сова | 120 | 23,64 |
| Линия яровой фиолетовой пшеницы | 541 | 106,56 |
| Сорт Памяти Азиева (кранозерная) | 129 | 25,41 |
| Линия яровой белозерной пшеницы(альбидум) | 66 | 13 |
Опыт 5
В течение трех лет нами были проведены анализы кормовых качеств сена сорта Сова. Анализ выполнялся в лаборатории качества кормов агрохимического центра «Омский»
Ниже представлены результаты качества сена сорта Сова за 3 года.
Таблица 11 – Качество сена сорта Сова
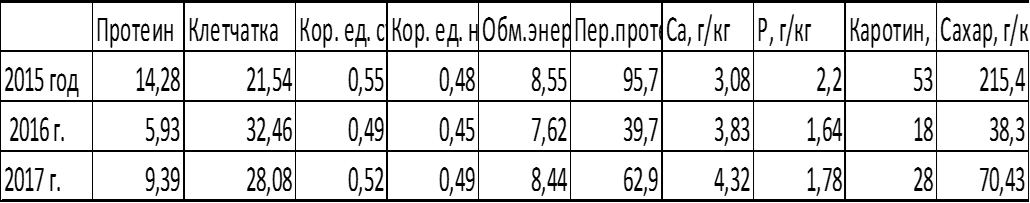
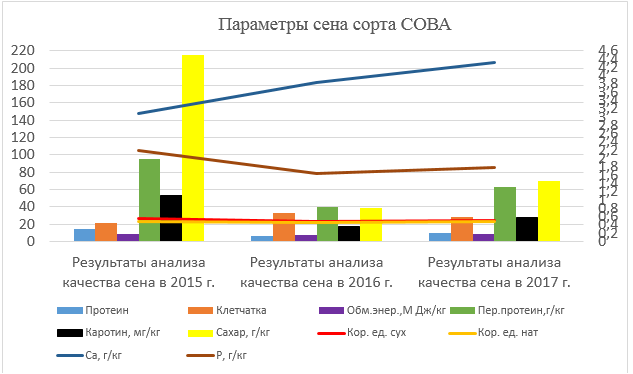
Рисунок 4 – Параметры сена сорта Сова
Исходя из графика можно сделать вывод, что наилучшим по качеству сена был урожай 2015 года. Далее наблюдалось ухудшение содержания сахара, перевариваемого протеина, каротина и протеина в 2016 и в 2017 году.
Опыт 6
Сканирование корней сорта Сова по годам жизни. Сканирование корней проводили в фазу выхода в трубку, отбор проводили в 3-х местах, на глубину 30 см. Сканирование корней проводилось с помощью программы Winrhizo. Данные представлены на рисунке 5.
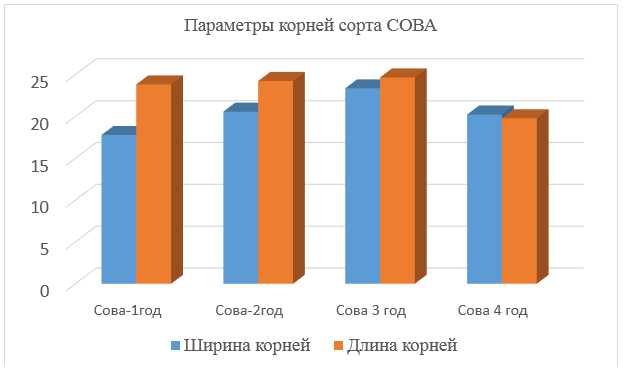
Рисунок 5 – Ширина и длинна корней сорта Сова
Максимальные показатели по ширине и длине корней достигаются на 3 год жизни, на 4 год жизни наблюдается ухудшение данных показателей.
Так же были определены такие параметры корневой системы как диаметр, объем и площадь корней растения пырея сизого сорта Сова.
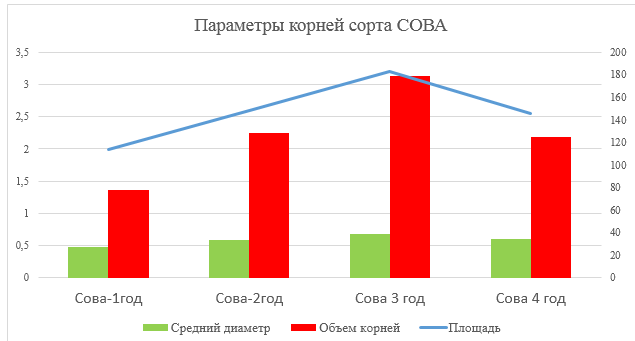
Рисунок 6 – Диаметр, объем и площадь корней сорта Сова
По показателям среднего диаметра, объема и площади корней наблюдается такая же тенденция что и с показателями ширины и длины корней.
Кроме того, были произведены измерения таких показателей, как число корневых кончиков, число корневых связей, число корневых перекрестков, число корневых вилок и суммарная длинна корней. Эти параметры представлены на рисунке 7.
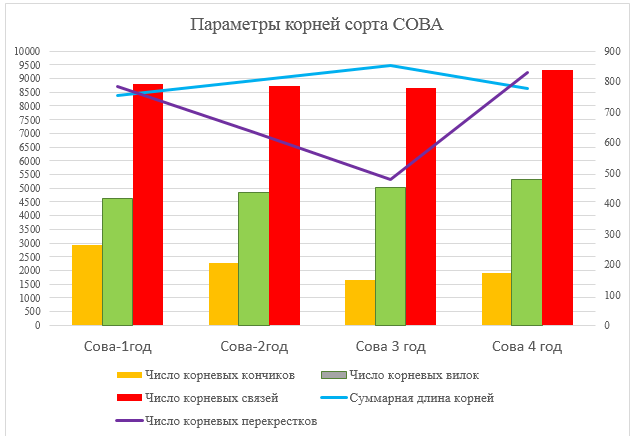
Рисунок 7 – Параметры корней сорта Сова
По числу корневых кончиков наблюдается ухудшение на 2 год жизни, число корневых связей остается на уровне, суммарная длина корней увеличивается до 3 года жизни.
На основе этих данных можно сделать вывод что, максимальное развитие корневой системы достигается на 3 год жизни, затем наблюдается ухудшение практически всех показателей корневой системы.
4.4 Зерна пырея сизого сорта Сова по содержанию белка и клейковины
Одним из важнейших показателей качества зерна является содержание белка и клейковины. Нами были проведены исследования данных параметров в течении трех лет, 2017-2019 гг. Результаты представлены в таблице 12.
Таблица 12 – Содержание белка и клейковины в зерне сорта Сова
| Питомник размножения | Производственный питомник размножения | ||
| 2017 | 2017 | ||
| белок | 16,2 | белок | 18,5 |
| клейковина | 34,4 | клейковина | 35,6 |
| 2018 | 2018 | ||
| белок | 16,3 | белок | 19,1 |
| клейковина | 34,7 | клейковина | 37,2 |
| 2019 | 2019 | ||
| белок | 17,3 | белок | 20,5 |
| клейковина | 34,7 | клейковина | 36 |
Исходя из данных можно сделать вывод, что качество зерна сорта Сова значительно выше яровой мягкой пшеницы. Качество белка сильно зависело от место расположения питомников. Содержание белка в питомнике у лесополосы значительно уступало содержанию белка в питомнике размножения, что обусловлено недостатком солнечного света.
Опыт 8
Изучение параметров зерновки сорта Сова в сравнении с пыреем диким и яровой мягкой пшеницей представлены в таблице 13.
Таблица 13 – Параметры зерновки сорта Сова
| Параметры | Сова урожай 2017 г | Сова урожай 2018 г | Сова урожай 2019 г | Пырей средний | Памяти Азиева 2017 | Памяти Азиева 2018 | Памяти Азиева 2019 | НСР 0,5 |
| Площадь, мм2 | 9,28 | 8,86 | 9,03 | 6,27 | 15,98 | 15,11 | 14,48 | 1,1 |
| Периметр, мм | 16,34 | 15,33 | 15,91 | 12,9 | 16,82 | 15,83 | 14,23 | 1,3 |
| Длина, мм | 7,02 | 6,60 | 6,83 | 5,52 | 6,47 | 6,21 | 6,45 | 0,55 |
| Ширина, мм | 1,68 | 1,67 | 1,67 | 1,44 | 3,29 | 3,30 | 3,00 | 0,81 |
| Циркулярность | 0,44 | 0,47 | 0,46 | 0,47 | 0,72 | 0,72 | 0,70 | 0,25 |
Из таблицы видно, что сорт Сова значительно превосходит по всем показателям зерновки пырей средний. Но также значительно уступает яровой мягкой пшенице. Параметры зерновки урожая 2018 года уступают параметрам зерновки урожая 2017 года, что обусловлено плохими погодными условиями в 2018 году.
4.5 Оценка влияния минеральных удобрений на сорт пырея сизого Сова
Объекты полевого опыта
Исследования проводили на опытном поле учебно-опытного хозяйства ФГБОУ ВО Омский ГАУ на лугово-черноземной среднемощной среднегумусовой тяжелосуглинистой почве. Объектом исследований являлись минеральные удобрения и пырей сизый сорт «Сова».
Полевой опыт был заложен в 2019 году по схеме:
- Без удобрений (контроль)
2. Подкормка в фазу кущения
3. Подкормка в фазу молочной спелости
4. Подкормка в фазу кущения и молочной спелости
5. N90
6. N90+ подкормка в фазу кущения
7. N90+ подкормка в фазу молочной спелости
8. N90+ подкормка в фазу кущения и молочной спелости
- N90P120
10. N90P120+ подкормка в фазу кущения
11. N90P120+ подкормка в фазу молочной спелости
12. N90P120+ подкормка в фазу кущения и молочной спелости
Повторность в опыте трехкратная с систематическим размещением вариантов. Общая площадь делянки 12 м2, учетная площадь 4,0 м2. Минеральные удобрения вносили весной после отрастания пырея сизого в форме карбамида и двойного суперфосфата вручную, подкормку проводили 10 % раствором мочевины.
Учет урожая проводили вручную методом пробного снопа 29 сентября в фазу полной спелости пырея сизого. По пробным снопам определяли структуру урожая: озернённость колоса, массу 1000 зёрен, долю зерна в общем биологическом урожае. Урожай зерна приводили к 100 %-ной чистоте и 14 %-ной стандартной влажности.
Сорт пырея сизого Сова получен в результате массового отбора зимостойких форм из американской популяции коллекционного образца пырея сизого.
Культура двойного направления на зерно и сено. Многолетний образ жизни. Хозяйственное использование до 7 лет.
Морфологические признаки: Ботаническое определение: Elytrigia intermedia (Host) Nevski subsp. Intermedia. Форма куста: полупрямостоячая, плотная. Стебли округлой формы, средней плотности, отсутствует опушение. Окраска узлов светлокоричневая. Сложный колос, узкий двусторонний, средней плотности. Окраска колоса сизо-зеленая. Остистость отсутствует. Колосковые чешуи – косообрубленные, цветочные чешуи – туповатые. Колоски продолговатой формы, окраска светло-зеленая. Язычок тупой короткий. Зерно светло-серое, фиолетовое. Ланцетной формы, бороздка не глубокая.
Хозяйственно-биологическая характеристика. Сова по вегетационному периоду относится к культурам позднеспелого типа. Продолжительность периода от всходов до полной спелости семян в конкурсном сортоиспытании варьировала от 110 до 120 суток. Сорт технологичен для возделывания, устойчив к полеганию и прорастанию зерна на корню.
Урожайность. Сорт Сова имеет очень хорошую урожайность семян для многолетних трав.
В конкурсном испытании по пару средняя урожайность за 2015‒2018 гг. составила 9,1 ц/га, что достоверно выше, чем у стандарта Ростовская 31 на 4,2 ц/га. Максимальная урожайность отмечена в 2018 г. на уровне 9,7 ц/га, стандарт Ростовская 31 соответственно 5,6 ц/га (достоверная прибавка составила 4,1 ц/га).
По урожайности зеленой массы и сена сорт Сова значительно превосходит стандарт. Так, средняя урожайность зеленой массы за годы испытания составила 259,3 ц/га, что достоверно выше стандарта Ростовская 31 на 58,8 ц/га. По показателям сена достоверная прибавка у сорта Сова составила 40,9 ц/га в сравнении со стандартом Ростовская 31 (103,2 ц/га и 62,3 ц/га соответственно).
Устойчивость к болезням. Обладает комплексной устойчивостью к болезням и вредителям.
Качество зерна. По качеству зерна сорт Сова имеет хорошие показатели по содержанию белка и клейковины. Средние показатели по качеству равны:
- содержание сырой клейковины 37,2 %,
- содержание сырого протеина 19,2 %,.
Сорт рекомендуется для степных и лесостепных зон регионов России.
Методы проведения исследований
Решение поставленных задач сопровождалось серией исследований и наблюдений, в работе используются методы полевых, лабораторных исследований, а также математический анализ с использованием ПЭВМ, которые позволяют объективно объяснить полученные результаты.
В период работ провели почвенную диагностику минерального питания пырея сизого, фенологические наблюдения, которые позволили установить сроки наступления фаз развития растений.
С целью контроля условий обеспеченности растений питательными веществами и коррекции питания использовали метод растительной (тканевой) диагностики минерального питания растений пырея сизого (фазы кущения и молочной спелости). На основании полученных данных провели некорневую подкормку растений пырея сизого в фазу кущения и молочной спелости 10-ным % раствором мочевины. Некорневая подкормка позволит устранить азотный дефицит в самом растении, а не в почве, удовлетворит потребности растений в азоте тогда, когда нельзя провести обычную подкормку или она малоэффективна. Использование некорневых подкормок иногда единственный прием из-за отсутствия удобрений в допосевной период.
В период работы над данной темой исследований проводились следующие анализы и наблюдения:
— определение запасов нитратного азота и продуктивной влаги в метровом слое почвы до посева и после уборки урожая;
— определение содержания в почве подвижных форм элементов минерального питания в диагностическом слое почвы 0-30 см;
— фенологические наблюдения за ростом и развитием растений по фазам;
— определение химического состава растений по фазам (кущение, молочная спелость, полная спелость). В растениях определяли валовое содержание азота, фосфора, калия и содержание минеральных форм этих элементов;
— учет урожая и определение его структуры (озерненность колоса, массу 1000 зёрен, долю зерна в общем биологическом урожае);
— определение качества зерна (содержание белка и клейковины).
Все лабораторные исследования проводили в учебно-научной лаборатории «Диагностика минерального питания и качества урожая сельскохозяйственных культур».
Определение содержания нитратного азота проводили по Грандваль-Ляжу, подвижного фосфора и обменного калия определяли по Чирикову. По всем фазам определение содержания валовых форм азота, фосфора и калия в растениях осуществляли из одной вытяжки, после мокрого озоления по Пиневич. Общий азот в полученном растворе определяли по Кьельдалю, фосфор по Дениже, калий на пламенном фотометре.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Перед внесением удобрений были отобраны почвенные образцы для определения в них доступных для растений элементов питания. Анализы показали, что содержание в почве нитратного азота находилось на очень низком уровне, подвижного фосфора высокое и обменного калия очень высокое (таблица 14).
Таблица 14– Содержание ЭМП в почве перед посевом, мг/кг
| Срок отбора | N- NО3 | Р2О5 | К2О |
| 15.05.2019 г. | 3,2 | 159 | 440 |
На основании данных содержания элементов питания и продуктивной влаги в почве был произведен расчет доз азотных и фосфорных удобрений.
5 Влияние минеральных удобрений и некорневых азотных подкормок на урожайность зерна пырея сизого
Перед отрастанием пырея сизого были отобраны почвенные образцы для определения продуктивной влаги в метровом слое почвы. Данные представлены в таблице 15.
Таблица 15 – Запасы продуктивной влаги в почве, 2019 г.
| Слой почвы, см | Влага | |
| % | мм | |
| 0-20 | 15,4 | 17,2 |
| 20-40 | 15,6 | 22,8 |
| 40-60 | 15,8 | 16,1 |
| 60-80 | 16,8 | 23,0 |
| 80-100 | 11,4 | 7,2 |
| 0-100 | 86,3 | |
Из данных таблицы 15 следует, что в условиях 2019 г. запасы продуктивной влаги в метровом слое почвы были на достаточно низком уровне и составляли 86,3 мм.
Исследования показали, что на варианте без удобрений урожайность пырея сизого составила 0,74 т/га (таблица 16).
| Таблица 16 ‒ Влияние расчетной дозы минеральных удобрений на урожайность зерна пырея сизого, 2019 г. |
| Вариант | Урожайность, т/га | Прибавка | |
| т/га | % | ||
| Без удобрений | 0,74 | — | — |
| N90 | 0,96 | 0,22 | 29,7 |
| N90P120 | 1,11 | 0,37 | 50,0 |
| НСР05 | 0,11 | 14,8 | |
Внесение азотных удобрений в дозе удобрений N90 и азотно-фосфорных в дозе N90P120 повысило урожайность зерна пырея сизого соответственно на 0,22 и 0,37 т/га.
Для формирования сильного зерна необходимо сбалансированное минеральное питание растений в течение всего вегетационного периода. В период вегетации ряд факторов внешней среды оказывает влияние на поступление питательных элементов из почвы в растение, в конечном итоге на формирование величины и качества урожая. Возникает ситуация несоответствия фактически сложившегося баланса ЭМП в растении тому оптимальному уровню, при котором формируется высокий биологически полноценный урожай.
Для контроля за минеральным питанием пырея сизого в фазы кущения и молочной спелости были отобраны растительные образцы для определения в них минеральных и валовых элементов питания. Данные химического анализа представлены в таблице 17.
Таблица 17 – Содержание минеральных элементов питания в растениях пырея сизого, 2019 г., мг%
| Вариант | Фаза развития растений | |||||
| кущение | колошение | |||||
| Nн | Рн | Кс | Nн | Рн | Кс | |
| Столыпинская 2 | ||||||
| Без удобрений | 42,4 | 50,5 | 980 | 37,5 | 28,5 | 825 |
| Подкормка (кущение) | 38,8 | 20,0 | 840 | |||
| N90 | 60,8 | 52,4 | 988 | 42,8 | 19,4 | 835 |
| N90 +подкормка | 38,5 | 19,0 | 760 | |||
| N90P120 | 61,2 | 52,5 | 960 | 40,4 | 21,2 | 790 |
| N90P120+подкормка | 45,4 | 22,2 | 760 | |||
| Оптимальный уровень | 60 | 50 | 920 | 37 | 19 | 720 |
| «в» | 0,28 | 0,26 | ||||
Данные таблицы 17 показывают, что в фазу кущения содержание неорганического азота в растениях пырея сизого на варианте без удобрений находилось ниже оптимального уровня, на удобренных вариантах – в пределах оптимального. Для оптимизации питания на варианте без удобрений на основе химического анализа растений, установленных оптимальных уровней элементов питания и коэффициента интенсивности («в») были рассчитаны дозы азотных для внесения в подкормку по формуле: Д = (Эо-Эф)2/в.Эо, где
Д – доза удобрения;
Эо – оптимальное содержание элемента;
Эф – фактическое содержание элемента;
в ‒ коэффициент интенсивности действия.
Рассчитанные дозы азота были внесены на варианте без удобрений в фазу кущения в виде некорневых подкормок 10- ным % раствором мочевины. На удобренных вариантах некорневая подкормка проводилась в дозе 30 кг/га д.в.
В фазу молочной спелости содержание неорганического азота в растениях пырея сизого находилось в пределах оптимального уровня. Несмотря на это в эту фазу нами была проведена некорневая азотная подкормка 10-% раствором мочевины в дозе 30 кг/га д.в.
Таблица 18 — Влияние некорневых азотных подкормок на урожайность зерна пырея сизого, 2019 г., т/га
| Фактор А,
подкормки |
Фактор В, удобрения | |||||
| без удобрений | N90 | N90P120 | ||||
| урожай
ность |
прибавка | урожай
ность |
прибавка | урожай
ность |
прибавка | |
| 1 | 0,74 | — | 0,96 | 0,22 | 1,11 | 0,37 |
| 2 (N15,2) | 0,86 | 0,12 | 1,00 | 0,04 | 1,12 | 0,01 |
| 3(N30) | 0,77 | 0,03 | 0,98 | 0,02 | 1,12 | 0,01 |
| 4(N15,2+ N30) | 0,88 | 0,14 | 1,02 | 0,06 | 1,13 | 0,02 |
НСР05, т/га, фактор А(удобрения) -0,11; фактор В (подкормки) – 0,08,
* 1) без подкормки, 2) подкормка в фазу кущения, 3) подкормка в фазу молочной спелости, 4) подкормка в фазу кущения + подкормка в фазу молочной спелости.
На варианте без удобрений применение некорневой подкормки в дозе 15,2 кг/га д.в. существенно повысило урожайность зерна пырея сизого. Прибавка урожая составила 0,12 т/га при НСР05 0,08 т/га. Урожайность без применения подкормки составила 0,74 т/га.
На удобренных вариантах применение некорневых азотных подкормок существенного влияния на урожайность пырея сизого не оказало. Это свидетельствует о том, что внесение минеральных удобрений в основное удобрение в полной мере обеспечило растения пырея сизого необходимыми элементами питания и вследствие этого дополнительного внесения удобрений в виде подкормок не требовалось.
Внесение некорневых азотных подкормок в фазу молочной спелости существенного влияния на урожайность изучаемого пырея сизого также не оказало.
5.1 Действие минеральных удобрений на качество зерна пырея сизого
Проведенные нами исследования показали, что применение некорневых азотных подкормок раствором мочевины в фазу кущения на показатели качества существенного влияния не оказали. В то же время применение некорневых азотных подкормок в фазу молочной спелости оказало положительное действие на качество зерна пырея сизого. Внесение 10-% раствора мочевины увеличило содержание белка с 15,7 до 16,4 и клейковины – с 30,8 до 32,1 %. При этом, все показатели качества зерна существенно выше на удобренных фонах. (таблица 19).
Таблица 19 – Показатели качества зерна пырея сизого, 2019 г., %
| Подкормки | Без удобрений | N90 | N90P120 | |||
| белок | клейковина | белок | клейковина | белок | клейковина | |
| 1 | 15,7 | 30,8 | 16,0 | 31,4 | 16,0 | 31,4 |
| 2 (N15,2) | 15,8 | 31,0 | 16,0 | 31,4 | 16,1 | 31,6 |
| 3(N30) | 16,1 | 31,7 | 16,2 | 31,7 | 16,2 | 31,8 |
| 4(N15,2+ N30) | 16,2 | 31,9 | 16,4 | 32,1 | 16,4 | 32,1 |
Следует отметить, что наибольшее содержание белка и клейковины в зерне пырея сизого было отмечено при применении некорневой подкормки в фазу кущения и молочной спелости на фоне основного удобрения N90P120.
5.2 Содержание элементов питания в растениях пырея сизого
Для определения основных агрохимических показателей нами был поведен химический анализ растений (зерно и солома) пырея сизого. Исследования показали, что содержание валовых азота, фосфора и калия в зерне составляли соответственно 2,80-2,94; 0,84-0,95 и 0,53-0,62 %, в соломе – 0,90-1,05; 0,30-0,40 и 1,03-1,08 % (таблица 20). При этом применение минеральных удобрений, как в основное внесение так и в подкормку, несколько увеличивало содержание элементов питания в растениях, в особенности азота.
Таблица 20 – Содержание валовых ЭМП в зерне пырея сизого, 2019 г., %
| Подкормки | Без удобрений | N90 | N90P120 | ||||||
| N | Р2О5 | К2О | N | Р2О5 | К2О | N | Р2О5 | К2О | |
| Зерно | |||||||||
| 1 | 2,80 | 0,87 | 0,53 | 2,85 | 0,86 | 0,54 | 2,86 | 0,90 | 0,55 |
| 2 (N15,2) | 2,82 | 0,88 | 0,55 | 2,86 | 0,84 | 0,58 | 2,87 | 0,92 | 0,58 |
| 3(N30) | 2,88 | 0,86 | 0,55 | 2,90 | 0,85 | 0,60 | 2,89 | 0,94 | 0,60 |
| 4(N15,2+ N30) | 2,90 | 0,88 | 0,58 | 2,94 | 0,86 | 0,62 | 2,93 | 0,95 | 0,62 |
| Солома | |||||||||
| 1 | 0,90 | 0,34 | 1,03 | 0,94 | 0,35 | 1,03 | 0,99 | 0,38 | 1,05 |
| 2 (N15,2) | 0,92 | 0,32 | 1,05 | 0,95 | 0,36 | 1,05 | 1,00 | 0,38 | 1,07 |
| 3(N30) | 0,95 | 0,30 | 1,03 | 0,97 | 0,36 | 1,06 | 1,03 | 0,37 | 1,06 |
| 4(N15,2+ N30) | 0,66 | 0,32 | 1,00 | 0,99 | 0,34 | 1,06 | 1,05 | 0,40 | 1,08 |
5.3 Вынос элементов питания растениями пырея сизого
Вынос питательных веществ с урожаем — важный показатель, который необходимо учитывать при определении потребности культур в удобрениях, расчете доз удобрений в конкретных условиях.
В таблице 21 приведены данные урожая сухого вещества зерна и соломы, в таблице 22 – выноса элементов питания зерном и соломой, в таблице 23 — общего выноса элементов питания (зерно + солома) растениями пырея сизого.
Таблица 21 – Урожайность сухого вещества пырея сизого, 2019 г. , т/га
| Подкормки | Удобрения | |||||
| без удобрений | N90 | N90P120 | ||||
| зерно | солома | зерно | солома | зерно | солома | |
| 1 | 0,64 | 5,18 | 0,83 | 7,10 | 0,96 | 8,21 |
| 2 (N15,2) | 0,74 | 6,19 | 0,86 | 7,52 | 0,96 | 8,40 |
| 3(N30) | 0,66 | 5,70 | 0,84 | 7,35 | 0,96 | 8,42 |
| 4(N15,2+ N30) | 0,76 | 6,51 | 0,88 | 7,55 | 0,97 | 8,46 |
Таблица 22 – Вынос валовых ЭМП растениями пырея сизого, 2019 г., кг/га
| Подкормки | Без удобрений | N90 | N90P120 | ||||||
| N | Р2О5 | К2О | N | Р2О5 | К2О | N | Р2О5 | К2О | |
| Зерно | |||||||||
| 1 | 17,9 | 5,57 | 3,39 | 23,6 | 7,13 | 4,48 | 27,7 | 8,64 | 5,28 |
| 2 (N15,2) | 20,9 | 6,51 | 4,07 | 24,6 | 7,22 | 4,99 | 27,6 | 8,83 | 5,57 |
| 3(N30) | 19,0 | 5,68 | 3,63 | 24,4 | 7,14 | 5,04 | 27,7 | 9,02 | 5,76 |
| 4(N15,2+ N30) | 22,0 | 6,69 | 4,41 | 25,9 | 7,57 | 5,46 | 28,4 | 9,22 | 6,01 |
| Солома | |||||||||
| 1 | 46,6 | 17,6 | 53,4 | 66,7 | 24,8 | 73,1 | 81,3 | 31,2 | 86,2 |
| 2 (N15,2) | 56,9 | 19,8 | 65,0 | 71,4 | 27,1 | 79,0 | 84,0 | 31,9 | 89,9 |
| 3(N30) | 54,2 | 17,1 | 58,7 | 71,3 | 26,5 | 77,9 | 86,7 | 31,2 | 89,2 |
| 4(N15,2+ N30) | 62,5 | 20,8 | 65,1 | 74,7 | 25,7 | 80,0 | 88,8 | 33,8 | 91,4 |
Таблица 23 – Общий вынос валовых ЭМП растениями пырея сизого, 2019 г.
| Подкормки | Без удобрений | N90 | N90P120 | ||||||
| N | Р2О5 | К2О | N | Р2О5 | К2О | N | Р2О5 | К2О | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
| Вынос, кг/га | |||||||||
| 1 | 64,5 | 23,2 | 56,8 | 90,3 | 31,9 | 77,6 | 109,0 | 39,8 | 91,5 |
| 2 (N15,2) | 77,8 | 26,3 | 69,1 | 96,0 | 34,3 | 84,0 | 111,6 | 40,7 | 95,5 |
| 3(N30) | 73,2 | 22,8 | 62,3 | 95,7 | 33,6 | 82,9 | 114,4 | 40,2 | 95,0 |
| 4(N15,2+ N30) | 84,5 | 27,5 | 69,5 | 100,6 | 33,3 | 85,5 | 117,2 | 43,0 | 97,1 |
| Потребность, кг/т | |||||||||
| 1 | 87,2 | 31,3 | 76,7 | 94,1 | 33,3 | 80,8 | 103,8 | 35,9 | 82,4 |
| 2 (N15,2) | 90,5 | 30,6 | 80,3 | 96,0 | 34,3 | 84,0 | 99,6 | 36,4 | 85,2 |
| 3(N30) | 95,1 | 29,6 | 80,9 | 97,6 | 34,3 | 84,6 | 102,1 | 35,9 | 84,8 |
| 4(N15,2+ N30) | 96,0 | 31,2 | 79,0 | 98,6 | 32,6 | 83,8 | 103,7 | 38,1 | 86,2 |
Из таблицы 23 видно, что для формирования 1 т зерна пырея сизого требуется 87,2-103,8 кг азота, 29,6-38,1 кг фосфора и 76,7-86,2 кг калия.
5.4 Коэффициенты использования яровой пшеницей элементов питания из почвы и удобрений
Коэффициенты использования элементов питания растениями из почвы обусловливаются биологическими особенностями культур, плодородием почвы, водным и тепловым режимами в течение вегетации и агротехникой возделывания. В таблице 24 представлены данные расчетов основных агрохимических показателей изучаемых сортов яровой пшеницы.
Таблица 24 — Коэффициент использования элементов питания из почвы растениями пырея сизого
| Агрохимические
показатели |
N | P2O5 | K2O |
| КИП,% | 54,9 | 6,1 | 5,4 |
| КИУ,% | 28,7 | 13,8 | — |
| Nм, кг/га | 58,7 |
Различные сельскохозяйственные культуры обладают неодинаковой способностью к усвоению отдельных питательных веществ из почвы и удобрений. Потребление растениями элементов питания в значительной мере зависит от их физиологического состояния и морфологического строения корневой системы.
Из данных таблицы следует, что КИП у пырея сизого составляет по азоту 54,9; по фосфору 6,1 и калию 5,4 %, КИУ по азоту 28,7; по фосфору 13,8 %. Азот мобилизации составил 58,7 кг/га.
Заключение
Наибольшая урожайность и прибавка урожая зерна пырея сизого в условиях 2019 г. была получена при внесении минеральных удобрений в основное внесение в дозе N90P120. Прибавка урожая зерна составила 0,37 т/га, при урожайности на варианте без удобрений 0,74 т/га.
Внесение некорневых азотных подкормок раствором мочевины на удобренных вариантах как в фазу кущения так и в фазу молочной спелости существенного влияния на урожайность зерна пырея сизого не оказало. Применение некорневых подкормок в фазу молочной спелости на всех вариантах существенно повысило качество зерна. В результате исследований определены основные (предварительные) агрохимические показатели для расчета доз минеральных удобрений на планируемый урожай и прибавку урожая изучаемых сортов яровой пшеницы:
— потребность элементов питания на формирование 1 т продукции составляет по азоту 87,2-103,8, по фосфору 29,6-38,1 и по калию 76,7-86,2 кг/т;
— КИП азота – 54,9; фосфора 6,1 и калия – 5,4;
— КИУ азота 28,7; фосфора – 13,8;
— Nм (азот мобилизации) – 58,7 кг/га.
6 Выделение высокоурожайных, устойчивых к болезням линий яровой мягкой пшеницы с фиолетовой окраской зерна
6.1 Изучение пшеницы с фиолетовым и голубым зерном в селекционном питомнике первого года
Для получения новых функциональных пищевых продуктов одним из актуальных направлений является селекция сортов пшеницы с повышенным содержанием биофлавоноидов – антоцианов, растительных соединений, способных оказывать положительное действие на здоровье человека.
Флавоноидные соединения являются естественными антиоксидантами (Prochazkova et al., 2011), их употребление в составе растительной пищи полезно для организма человека и благотворно влияет на различные патологические состояния, включая сердечно-сосудистые заболевания, артриты, различные виды рака и болезнь Альцгеймера (Pascual-Teresa et al., 2010; Simoes, 2012; Pojer et al., 2013). Данные соединения также полезны в лечении сахарного диабета 2 типа и ожирения (Yawadio et al., 2007; Prior et al.,2008).
Синтез различных окрашенных флавоноидных соединений в определенных структурах зерна злаковых растений приводит к появлению окраски. В результате синтеза антоцианов семена злаков могут приобретать окраску различных оттенков от голубовато-серого и красноватого до темно-фиолетового и почти черного. Среди различных соединений, придающих окраску зерну злаковых растений, наибольшим антиоксидантным потенциалом обладают антоцианы (Knievel et al., 2009). Антоцианы могут накапливаться в различных частях растения. Их основная физиологическая роль заключается в защите растений от избыточного УФ-излучения, а так же содержание антоцианов возрастает при неблагоприятном воздействии окружающей среды (засуха, холод, засоление и т.д.;) (Chalker-Scott, 1999).
Основная задача исследований направлена на создание высокоурожайных сортов пшеницы с фиолетовой и голубой окраской зерна, с высоким содержанием биофлавоноидов, а также устойчивостью к биотическим и абиотическим факторам среды. Особое значение при этом имеет получение качественной зерновой продукции с высокими хлебопекарными и органолептическими показателями для изготовления хлебобулочных изделий отличающихся антиоксидантной активностью.
В питомнике первого года испытаний (СП1) в 2019 г. высеяно 786 линий с фиолетовой и голубой окраской перикарпа пшеницы. Линии представляли собой потомство одного колоса, выбранного в результате индивидуальных отборов в 2018 г, а также новые линии фиолетовой пшеницы, полученные при сотрудничестве с ИЦиГ СО РАН и СИММИТ. Посев проводился вручную 17 мая. В качестве стандартов были посеяны сорта – Памяти Азиева, Дуэт, Элемент 22.
В ходе исследований проводились оценки устойчивости линий к грибным патогенам (мучнистая роса – Erysiphe (Blumeria) graminis, бурая ржавчина – Puccinia triticina, стеблевая ржавчина – Puccinia graminis), фенологические наблюдения, а также определено качество и урожайность зерна.
В результате изучения, было отобрано 200 линий для дальнейшего изучения в селекционном питомнике второго года (СП2), включения в питомник гибридизации и 40 колосьев для посева в СП1 в 2020 г.
Все отобранные линии по вегетационному периоду относятся к среднепозднему типу, устойчивы к бурой и стеблевой ржавчине пшеницы, устойчивы к полеганию. Степень устойчивости линий к мучнистой росе варьируется по образцам.
Линии в таблице BC1F4 поколения – гомозиготы по генам антоциановой окраски – с темно фиолетовым зерном (интогрессии в двух хромосомах: 2А и 7D) и голубым зерном (замещение пырейной хромосомой 4Ag / 4B или 4D хромосомы мягкой пшеницы).
Таблица 25 – Результаты изучения линий фиолетовой пшеницы отобранных в питомнике СП1.
| Линии поколения BC1F4 | Цвет зерна | Дата коло-шения | Оценка устойчивости к болезням, степень поражения / тип поражение | Количе-ство
отобран-ных линий, шт. |
||
| Муч-нистая роса, балл | Стеблевая ржавчина | Бурая ржав-чина | ||||
| 2-Элемент x i:S29 PF | фиолетовое | 18 июля | 3 | 20M | 10MS | 10 |
| 2-2-Элемент x i:S29 PF | фиолетовое | 19 июля | 5 | 20M | 15M | 20 |
| 2-3-Элемент x i:S29 PF | фиолетовое | 19 июля | 5 | 20M | 5M | 10 |
| 2-4-Элемент x i:S29 PF | фиолетовое | 19 июля | 5 | 20M | 10MS | 10 |
| 2-5-Элемент x i:S29 PF | фиолетовое | 20 июля | 5 | 20M | 10MS | 10 |
| 3-s:S29_4Ag (Ba)/4D x Элемент 22 | голубое | 20 июля | 7 | 20M | 5M | 10 |
| 3-1-s:S29_4Ag (Ba)/4D x Элемент 22 | голубое | 21 июля | 9 | 20M | 5M | 10 |
| 3-2-s:S29_4Ag (Ba)/4D x Элемент 22 | голубое | 20 июля | 7 | 20M | 10MS | 10 |
| 4-s:S29_4Ag (Ba)/4D x Элемент 22 | голубое | 21 июля | 9 | 10MS | 10MS | 10 |
| 4-1-s:S29_4Ag (Ba)/4D x Элемент 22 | голубое | 21 июля | 7 | 20M | 10MS | 10 |
| 4-2-s:S29_4Ag (Ba)/4D x Элемент 22 | голубое | 21 июля | 9 | 15M | 20MS | 10 |
| 10-Турецкая x i:S29 PF | фиолетовое | 19 июля | 1 | 20MS | 20MS | 5 |
| 10-2-Турецкая x i:S29 PF | фиолетовое | 18 июля | 1 | 10MS | 10MS | 5 |
| 10-3-Турецкая x i:S29 PF | фиолетовое | 18 июля | 1 | 20MS | 20MS | 5 |
| 10-4-Турецкая x i:S29 PF | фиолетовое | 18 июля | 7 | 20MS | 20MS | 5 |
| 11-Турецкая x s:S29_4Ag (Ba)/ 4D | голубое | 20 июля | 7 | 40MS | 40MS | 10 |
| 11-1-Турецкая x s:S29_4Ag (Ba)/ 4D | голубое | 18 июля | 7 | 20MS | 20MS | 5 |
| 11-3-Турецкая x s:S29_4Ag (Ba)/ 4D | голубое | 18 июля | 9 | 30MS | 30MS | 5 |
| 11-4-Турецкая x s:S29_4Ag (Ba)/ 4D | голубое | 22 июля | 9 | 10MS | 10MS | 5 |
| 11-5-Турецкая x s:S29_4Ag (Ba)/ 4D | голубое | 20 июля | 9 | 10M | 10MS | 5 |
| 11-6-Турецкая x s:S29_4Ag (Ba)/ 4D | голубое | 19 июля | 9 | 10M | 10M | 10 |
| Памяти Азиева | красное | 12 | 5 | 90S | 30S | — |
| Дуэт | красное | 15 | 7 | 90S | 30S | — |
| Элемент 22 | красное | 19 | 7 | 10M | 5M | — |
Расшифровка линий-доноров антоциановой окраски:
1. Донор фиолетовой окраски — изогенная линия i:S29 Pp3Pp1PF (сокращение – i:S29 PF) на основе сорта Саратовская 29 с двумя интогрессивными вставками: в длинное плечо 2A (с доминантным аллелем гена Pp3) и в короткое плечо 7D хромосомы (с доминантным аллелем гена Pp1) от сорта Purple Feed (PF) с фиолетовым перикарпом зерна.
2. Донор голубой окраски — замещенная пшенично-пырейная линия s: S29_4Ag (Ba)/ 4D на основе сорта Саратовская 29 с замещенной целой хромосомой от пырея высокого Agropyron elongatum Host (decaploid tall wheatgrass; 2n = 10x = 70, StStStStEeEeEbEbExEx; syn. Thinopyrum ponticum; Elytrigia pontica).
7 Изучение коллекции полбы в селекционном питомнике первого года
Полба – ценная сельскохозяйственная культура (Triticum dicoccum, спельта, двузернянка, эммер), неприхотливое, скороспелое яровое растение, обладающее широкой экологической пластичностью (Воробейков, Кондрат, 2007). В древности полба была одной из самых распространенных пшениц. В настоящее время этот хлебный злак активно культивируют на небольших площадях в Центральной Европе, Дагестане, Белгородской, Ярославской областях, районах Среднего Поволжья, Предуралье, Сибири. Ценной биологической особенностью полбы является ее скороспелость (Юков, 2005).
Благодаря хорошо развитой корневой системе, полба отличается засухоустойчивостью, в засушливые годы она дает более высокие урожаи, чем другие зерновые культуры, в том числе на бедных почвах (Артюшенко, 1973). Полба менее других зерновых культур повреждается болезнями и вредителями (Тюнин и др., 2017), значительно превосходит пшеницу по содержанию белка (от 25 до 37 %), насыщенных жирных кислот, клетчатки, железа, витаминов группы В, отличается низким содержанием клейковины и глютена (Муслимов, Исмаилов, 2012; Темирбекова и др., 2010; Яркова, Якунина, 2018). Колос полбы отличается невысокой урожайностью и повышенной ломкостью, что приводит к большим потерям зерна при уборке урожая (Смирнова и др., 1989). Существенный недостаток этой культуры – трудность ее обмолачивания и последующая технологическая обработка.
В связи с этими недостатками, большой интерес представляют голозерные высокоурожайные сорта полбы.
В 2019 г. в питомнике СП1 было высеяно 12 гибридных линий F0. Все полученные линии первого поколения (F1) были отобраны для дальнейшего селекционного процесса и передачи в питомник второго года исследований.
Таблица 26 – Линии полбы поколения F1 для передачи в СП2.
| № п/п | Линии поколения F1 |
| 1 | Полба 1/ к-6436 |
| 2 | Полба 1 / к-7502 |
| 3 | Полба 1 / к-11750 |
| 4 | Полба 1 / к-13085 |
| 5 | Полба 1 / к-21582 |
| 6 | Полба 1 / к-23647 |
| 7 | Полба 1 / к-23649 |
| 8 | Полба 1 / к-23652 |
| 9 | Полба 1 / к-35890 |
| 10 | Полба 1 / к-37396 |
| 11 | Полба 1 / к-40170 |
| 12 | Полба 1 / к-64408 |
В питомнике гибридизации в 2019 г. получено 7 гибридных комбинаций F0 (250 зерен) от скрещивания сортов Полба 1 и Полба Греммэ с сортами полбы из коллекции, передано в СП1 2020 г.
8 Изучение линий фиолетовозерной пшеницы в селекционном питомнике второго года
В 2019 г. на опытном поле Омского ГАУ был заложен опыт по изучению фиолетовозерной пшеницы. Материал был получен в 2018 году из ИЦиГ СО РАН. Изучаемый материал состоял из 79 линий. Линии изучались на делянках площадью 1 м2. Посев проводился 17 мая, сеялкой ССФК-7. Предшественник – чистый пар. Норма высева 500 всх. зерен на 1м2. Через каждые 20 номеров в качестве стандартов высевали сорта: Памяти Азиева, Дуэт и Элемент 22.
Оценки, учеты и наблюдения проводили в соответствии с методическими указаниями ВИР по изучению пшеницы. Математическая обработка данных по урожайности проведена методом дисперсионного анализа (Доспехов, 1985). Тип поражения оценивался по шкале Майнса и Джексона с международной буквенной системой обозначения: R – устойчивый, TR- прослеживается устойчивость, MR – умеренно стойкий, MS – умеренно восприимчивый, M – перекрывание (MR и MS, присутствуют два вида пустул), MSS – умеренно восприимчивый к восприимчивому, TS – прослеживается восприимчивость, S – восприимчивый.
Для развития стеблевой и бурой ржавчины сложились благоприятные метеорологические условия, инфекционный фон был неравномерный, изучаемые образцы поразились в низкой либо в средней степени, либо признаки поражения отсутствовали.
В течение вегетационного периода проводили фенологические наблюдения, полевая оценка, оценка на устойчивость к бурой и стеблевой ржавчине. По результатам оценок и наблюдений отобрано 19 линий. Стандарты и отобранные номера убирали в фазу полной восковой спелости вручную (серпом). Селекционный материал обмолачивался сноповой молотилкой МПСУ-500. Для определения урожайности сухое зерно взвешивали.
В таблице 27 представлены данные по урожайности и вегетационному периоду лучших линий в СП2 (1 м2).
Таблица 27 – Урожайность и продолжительность вегетационного периода лучших линий СП2 (1 м2)
| Сел. номер | Сорт, образец | Масса зерна, г/м2 | Превышение стандарта,
± г/м2 |
Вегетационный период, суток |
| 1 | 2 | 3 | 4 | 5 |
| Среднеранние | ||||
| Памяти Азиева, стандарт | 374 | 72 | ||
| 285 | BC1F2 (Турецкая х BC5F2) | 403 | +29 | 72 |
| 281 | BC1F2 (Турецкая x i:S29 PF) | 417 | +43 | 76 |
| 286 | BC1F2 (Турецкая х BC5F2) | 419 | +45 | 76 |
Окончание таблицы 27
| 1 | 2 | 3 | 4 | 5 |
| 275 | BC1F2 (Турецкая x i:S29 PF) | 440 | +66 | 72 |
| Среднеспелые | ||||
| Дуэт, стандарт | 380 | 78 | ||
| 301 | BC1F2 (Айна х i:S29 P) | 417 | +37 | 78 |
| 298 | BC1F2 (Айна х i:S29 PF) / (i:S29 PF x Айна) | 461 | +81 | 79 |
| 297 | BC1F2 (Айна х i:S29 PF) / (i:S29 PF x Айна) | 522 | +142 | 79 |
| Среднепоздние | ||||
| Элемент 22, стандарт | 490 | 85 | ||
| 240 | BC1F2 (Элемент x I:S29 PF | 522 | +32 | 84 |
| 250 | BC1F2 (Элемент x I:S29 PF | 539 | +49 | 86 |
| 264 | BC1F2 (Элемент x I:S29 PF | 546 | +56 | 84 |
| 267 | BC1F2 (Элемент x I:S29 PF | 548 | +58 | 85 |
| 366 | BC1F2 (Элемент x I:S29 PF | 570 | +80 | 85 |
| 251 | BC1F2 (Элемент x I:S29 PF | 584 | +94 | 86 |
| 253 | BC1F2 (Элемент x I:S29 PF | 593 | +103 | 85 |
| 255 | BC1F2 (Элемент x I:S29 PF | 597 | +107 | 84 |
| 249 | BC1F2 (Элемент x I:S29 PF | 617 | +127 | 86 |
| 254 | BC1F2 (Элемент x I:S29 PF | 647 | +157 | 85 |
| 260 | BC1F2 (Элемент x I:S29 PF | 650 | +160 | 84 |
| 239 | BC1F2 (Элемент x I:S29 PF | 687 | +197 | 84 |
| НСР0,05 | 28,5 | |||
В группе среднеранних линий 4 лучших образца превосходили стандарт Памяти Азиева по урожайности. Номера превышают по урожайности стандарт (от +29 до +66 г/м2).
В среднеспелой группе по урожайности выделились 3 образца № 301 BC1F2 (Айна х i:S29 P) – 417 г/м2, № 298 BC1F2 (Айна х i:S29 PF) / (i:S29 PFx Айна) – 461 г/м2, № 297 BC1F2 (Айна х i:S29 PF) / (i:S29 PF x Айна) – 522 г/м2. Вегетационный период находился в пределах 78 ‒ 79 суток.
В среднепоздней группе 12 образцов превзошли стандарт Элемент 22. Вегетационный период находился в пределах 84 – 86 суток.
Из 79 изучаемых образцов 19 переданы в контрольный питомник.
9 Изучение линий фиолетовозерной пшеницы и полбы в конкурсном сортоиспытании
Цель исследований — на основе проведения в условиях полевого опыта сравнительной оценки новых сортов яровой мягкой пшеницы по важнейшим хозяйственно-полезным признакам и свойствам выявить лучшие из них, существенно превосходящие соответствующие стандарты по урожайности, качеству зерна и другим показателям, которые учитываются при включении сорта в Государственный реестр селекционных достижений, допущенных к использованию.
Материал и методика проведения исследований
В опытах изучали лучшие сорта, выделенные в контрольном питомнике, предварительном и конкурсном сортоиспытаниях в 2016-2018 г.г.
В качестве стандартов использовали среднеранний сорт Памяти Азиева, среднеспелый — Дуэт и среднепоздний – Элемент 22.
Посев проводили 27 мая. Конкурсное сортоиспытание (КСИ) закладывали по чёрному пару и второй культурой после пара. Коэффициент высева 4,5. Учётная площадь делянки 25 м2. Повторность в КСИ по пару — четырёхкратная, второй культурой после пара – трёхкратная.
Все наблюдения, учёты, оценки и анализы проводили по методике Госсортсети. Уборка урожая однофазная комбайном «Сампо 130». Данные об урожайности зерна изучаемых сортов обрабатывали методом дисперсионного анализа по Б.А. Доспехову. Определение содержания белка и клейковины проводили на приборе «Инфралюм» ФТ 10.
В конкурсном сортоиспытании по пару изучали 19 образцов яровой мягкой пшеницы и 2 образца полбы. По вегетационному периоду, 8 образцов относятся к среднеранней группе, 8 образцов к среднеспелой группе и 5 образцов к среднепоздней группе. В группе среднеранних образцов, все образцы показали лучшую устойчивость к мучнистой росе, чем стандарт Памяти Азиева. Устойчивость в 4 балла показали образцы Эритроспермум 4-16 и Полба Руно, более высокую устойчивость в 5 баллов отмечено у образцов – Лютесценс 13-15 и Полба 1. В слабой степени, стеблевой ржавчиной поразились Лютесценс 53-15 (10М), Лютесценс 13-15 (5М), Эритроспермум 4-16 (5М), Лютесценс 34-16 (10М), Лютесценс 63-16 (5М) и показали полную устойчивость к бурой ржавчине.
Таблица 28 — Характеристика сортов пшеницы в конкурсном сортоиспытании по пару
| № п/п | Сорт | Вегетационный период, сут, | Урожайность, т/га | Поражение стеблевой ржавчиной | Поражение бурой ржавчиной | Поражение мучнистой росой, балл | Общая селекционная оценка | Содержание клейковины, % | Содержание белка, % |
| 1 | Памяти Азиева (st) | 82 | 2,99 | 80S | 50S | 2 | 3,19 | 23,57 | 14,16 |
| 2 | Лютесценс 53-15 | 83 | 4,14 | 10M | R | 3 | 3,5 | 30,21 | 16,56 |
| 3 | Лютесценс 13-15 | 82 | 4,82 | 5M | R | 5 | 3,81 | 28,44 | 15,45 |
| 4 | Эритроспермум 4-16 | 82 | 4,83 | 5M | R | 4 | 4,06 | 29,09 | 15,32 |
| 5 | Лютесценс 34-16 | 82 | 4,87 | 10M | R | 2 | 3,5 | 23,30 | 13,40 |
| 6 | Лютесценс 63-16 | 83 | 4,38 | 5M | R | 2 | 3,75 | 27,78 | 15,07 |
| 7 | Полба 1 | 76 | 2,85 | 50S | 0 | 5 | 3,63 | 26,17 | 14,87 |
| 8 | Полба Руно | 76 | 2,99 | 50S | 0 | 4 | 2,94 | — | — |
| 9 | Дуэт (st) | 85 | 2,99 | 80S | 40S | 4 | 3,25 | 24,53 | 13,94 |
| 10 | Лютесценс 123-13 | 85 | 3,47 | 20M | 5M | 5 | 3,5 | 30,69 | 16,09 |
| 11 | Лютесценс 126-15 | 85 | 4,22 | 10M | R | 4 | 4,38 | 26,98 | 14,43 |
| 12 | Эритроспермум 96-15 | 85 | 4,43 | 10M | 5M | 3 | 4,63 | 26,95 | 14,25 |
| 13 | Лютесценс 89-15 | 83 | 4,76 | 10M | R | 3 | 3,5 | 28,15 | 15,01 |
| 14 | Лютесценс 66-16 | 85 | 4,25 | 10M | R | 3 | 3,81 | 27,71 | 15,23 |
| 15 | Фиолетовая Purple2-2018 | 78 | 3,00 | 80S | 50S | 2 | 2,56 | 20,50 | 14,15 |
| 16 | Фиолетовая Perple5-2018 | 78 | 2,88 | 100S | 100S | 1 | 3 | 20,97 | 13,81 |
| 17 | Элемент 22 (st) | 90 | 3,98 | 5M | R | 5 | 4,13 | 30,10 | 15,57 |
| 18 | Лютесценс 88-13 | 86 | 5,02 | 5M | R | 4 | 4,38 | 26,28 | 14,86 |
| 19 | Лютесценс 19-16 | 91 | 4,19 | 10M | R | 4 | 3,75 | 26,60 | 14,14 |
| 20 | Эритроспермум 5-17 | 83 | 4,66 | R | R | 4 | 4,13 | 26,18 | 14,83 |
| 21 | Лютесценс 34-17 | 83 | 5,03 | 10M | 20MS | 1 | 4,06 | 27,94 | 14,02 |
| НСР 0,5 | 0,36 |
В группе среднеспелых образцов, куда относиться 6 образцов, по устойчивости к мучнистой росе можно выделить Лютесценс 123-13 (5 баллов) и на уровне стандарта в 4 балла Лютесценс 126-15. По устойчивости к стеблевой ржавчине, все образцы поражались в средней мере, по устойчивости бурой ржавчиной выделяются образцы Лютесценс 126-15, Лютесценс 89-15, Лютесценс 66-16. Группа среднепоздних сортов представлена 5 образцами. По устойчивости к мучнистой росе ни один образец не смог превзойти стандарт Элемент 22 равной 5 баллов. Практически у всех образцов поражение бурой ржавчиной на уровне стандарта, полную устойчивость к стеблевой ржавчине показал образец Эритроспермум 5-17, остальные образцы, оказались пораженными в слабой степени (5-10М).
Два образца фиолетовозерной пшеницы показали урожайность на уровне сорта Памяти Азиева из-за гораздо более высокого поражения бурой и стеблевой ржавчиной. Так же поражение болезнями достоверно снизило содержание в зерне белка и клейковины. Рекомендуется продолжить изучение этих образцов при обработке фунгицидами, а так же использовать их в качестве исходного материала для гибридизации в 2020 году, с целью получения на их базе устойчивых к болезням сортов.
10 Рыночные перспективы новых сортов селекции Омского ГАУ
В Омском ГАУ на сегодняшний день ведется работа над созданием уникальных в рыночном отношении сортов пшеницы и полбы. Среди сортов пшеницы особенно выделяются фиолетовозерная пшеница и пырей сизый (многолетняя культура двойного назначения – зерно и сено), созданная на основе диких злаков.
Рыночные перспективы пырея сизого связаны с относительно высокой урожайностью зерна и сена, а также продолжительным сроком жизни – до 7 лет. Кроме того, технология возделывания сорта предполагает использование гербицидов только первые два года.
В таблице 29 представлен расчет экономической эффективности возделывания пырея сизого по сравнению с традиционной пшеницей.
На первый взгляд сорт пырея сизого существенно уступает традиционным сортам пшеницы в урожайности, а более низкая масса 1000 семян делает его еще менее конкурентоспособным на рынке. При этом качество зерна пырея сизого позволяет использовать его не только на кормовые, но и на пищевые цели. Кроме того, существенным рыночным преимуществом данного сорта является наличие еще одного вида товарной продукции, которая недоступна традиционным сортам яровой мягкой пшеницы – значительная урожайность сена высокого качества. Еще одним неоспоримым преимуществом пырея сизого является его продолжительный срок жизни, что уже со второго года практически на половину сокращает производственные расходы на 1 га. Дополнительным преимуществом пырея сизого является потребность в гербицидах только первые два года, в то время как традиционные сорта яровой мягкой пшеницы требуют внесения гербицидов ежегодно.
Имеющиеся преимущества пырея сизого в конечном итоге в расчете на 7 лет позволяют получить совокупный доход с одного га в размере более 398 тыс. руб., что в 2,7 раза больше по сравнению с доходами с одного га у традиционных для Омской области сортов яровой мягкой пшеницы, площадь посева под которыми составляет около 60% всей посевной площади яровой мягкой пшеницы в регионе. Расчетная рентабельность продаж за 7 лет по сорту пырея сизого составляет 92,66%, в то время как по традиционным сортам пшеницы не превышает 64%.
Таблица 29 – Сравнительный анализ экономической эффективности возделывания пырея сизого по сравнению с традиционными сортами пшеницы в условиях Омской области
| Показатель | Пырей сизый | Стандартный сорт яровой мягкой пшеницы |
| Срок жизни, лет | 7 | 1 |
| Расходы на гербициды 1 год, руб./га | 600,0 | 600,0 |
| Расходы на гербициды 2 год, руб./га | 600,0 | 600,0 |
| Расходы на гербициды 3-7 год, руб./га | 0,0 | 600,0 |
| Итого расходы на гербициды за 7 лет, руб./га | 1 200,0 | 4 200,0 |
| Производственные расходы на 1 га (посев, уход, уборка) — первый год | 7 000,0 | 7 000,0 |
| Производственные расходы на 1 га — второй и последующие годы | 3 500,0 | 7 000,0 |
| Итого производственные расходы на 1 га за 7 лет, руб. | 29 200,0 | 53 200,0 |
| Урожайность зерна, ц/га | 9,1 | 30,0 |
| Итого валовой сбор зерна с 1 га за 7 лет, ц | 63,7 | 210,0 |
| Урожайность сена с 1 га, ц | 259,3 | 0,0 |
| Итого валовой сбор сена с 1 га за 7 лет, ц | 1 815,1 | 0,0 |
| Затраты на 1 ц зерна (40% побочная продукция) | 275,0 | 253,3 |
| Затраты на 1 ц сена, руб. | 6,4 | 0,0 |
| Стоимость 1 ц зерна, руб. | 550,0 | 700,0 |
| Доходы от реализации зерна за 7 лет, руб. | 35 035,0 | 147 000,0 |
| Доходы от реализации сена за 7 лет, руб. | 363 020,0 | 0,0 |
| Итого доходы за 7 лет с 1 га, руб. | 398 055,0 | 147 000,0 |
| Итого рентабельность продаж за 7 лет, % | 92,66 | 63,81 |
*производственные расходы на 1 га взяты исходя из суммы средних расходов по учебно-опытному полю Омского ГАУ, цены реализации – среднерыночные по Омской области в 2019 г.
Рыночные перспективы сортов фиолетовозерной пшеницы связаны с повышенным содержанием антиоксидантов и микроэлементов. При этом в конкурсном испытании данные сорта показали определенные недостатки, связанные, прежде всего, с высоким уровнем поражения бурой и стеблевой ржавчиной. Не смотря на это, урожайность сортов фиолетовозерной пшеницы осталась на уровне стандарта, что указывает на высокий рыночный потенциал при применении фунгицидов. Дальнейшие исследования в данном направлении позволят Омскому ГАУ занять пока свободную в России нишу сортов яровой мягкой пшеницы с фиолетовой окраской зерна. Данные сорта имеют высокий потенциал на рынке функционального питания, как и сорта полбы.
В конкурсном сортоиспытании сорта полбы показали высокую устойчивость к поражению бурой ржавчиной и мучнистой росой, продемонстрировав при этом урожайность на уровне стандарта. С экономической точки зрения это означает отсутствие у предприятий, возделывающих данные сорта, расходов на средства защиты от болезней. В среднем в условиях Омской области такие расходы составляют около 600 руб. на 1 га. Таким образом, уже сейчас по сравнению со стандартом сорта полбы демонстрируют повышенную экономическую эффективность: расходы на 1 га ниже на 7,9%, рентабельность продаж выше на 3 п.п. (таблица 30)
Таблица 30 – Сравнительная экономическая эффективность возделывания полбы и яровой мягкой пшеницы в условиях Омской области
| Показатель | Полба Руно | st Памяти Азиева |
| Производственные расходы на 1 га всего, руб., в т.ч. | 7 000,0 | 7 600,0 |
| расходы на гербициды, руб./га | 0,0 | 600,0 |
| Урожайность, т/га | 3,0 | 3,0 |
| Цена реализации 1 тонны, руб. | 7 000,0 | 7 000,0 |
| Валовая выручка, руб./га | 20 930,0 | 20 930,0 |
| Прибыль от реализации, руб./га | 13 930,0 | 13 330,0 |
| Рентабельность продаж, % | 66,56 | 63,69 |
*производственные расходы на 1 га взяты исходя из суммы средних расходов по учебно-опытному полю Омского ГАУ, цены реализации – среднерыночные по Омской области в 2019 г.
Таким образом, новые сорта селекции Омского ГАУ имеют высокий рыночный потенциал и способны сформировать новые рыночные ниши, в том числе в сфере органического земледелия и функционального питания.
Заключение
Комплексная оценка линий многолетней пшеницы, полученных скрещиванием мягкой пшеницы с различными видами пырея
- В результате проведенных полевых и структурных анализов а также анализа на качество зерна, было выделено 5 лучших образцов: № 2 (Agrotana), № 4 (119550), №10 (235а), №11 (TAF46), №12 (Ot 38).
- Выделявшиеся образцы были пересеяны в питомник в 3-х кратной повторности делянками с площадью 1м для дальнейшего изучения.
Изучение доместицированного пырея сизого
- Оптимальной площадью питания для растения являются 40 х 20 и 20 х 10, но экономически более эффективным является посев с площадью питания 40 х 20.
- Положительный эффект длины растения был на признаках длины колоса, массы растения, массы стебля и числа колосков с колоса. Высокорослые растения сорта Сова достоверно превосходили низкорослые по этим показателям.
- В результате проведения опыта установлено, что оптимальным сроком укоса сорта Сова на сено является 1 декада августа.
- Максимальное развитие корневой системы достигается на 3-ий год жизни, затем наблюдается ухудшение практически всех показателей корневой системы.
- Качество зерна сорта Сова значительно выше яровой мягкой пшеницы. Качество белка сильно зависело от место расположения питомников. Содержание белка в питомнике у лесополосы значительно уступало содержанию белка в питомнике размножения, что обусловлено недостатком солнечного света.
- Сорт Сова значительно превосходит по всем показателям зерновки пырей средний. Но также значительно уступает яровой мягкой пшенице. Параметры зерновки урожая 2018 года уступают параметрам зерновки урожая 2017 года, что обусловлено плохими погодными условиями в 2018 году.
Выделение высокоурожайных, устойчивых к болезням линий яровой мягкой пшеницы с фиолетовой окраской зерна
- В 2019 году в селекционном питомнике второго года было изучено 79 новых образцов фиолетовозерной пшеницы, 19 лучших образцов переданы в контрольный питомник.
- В конкурсном сортоиспытании было изучено два образца фиолетовозерной пшеницы, показавших урожайность на уровне стандарта. Рекомендовано продолжить их изучение в селекционном процессе, а так же использовать в качестве исходного материала для гибридизации.
Список используемых источников
- AACC, 2000. American Association Of Cereal Chemists International. Approved Method No: 10-10B: St. Paul. MN., USA.
- AACCI, 2010. American Association Of Cereal Chemists International. Approved Method No: 32-07.01, 46-30.01, 54–21.02, 74-09.01, The Association: St. Paul. MN., USA.
- Abbo S. Pinhasi van-Oss R, Gopher A, Saranga Y, Ofner I, Peleg Z. Plantdomestication versus crop evolution: a conceptual framework for cereals andgrain legumes. Trends Plant Sci 2014;19(6):351–60.
- Abdel-Aal E.-S.; Hucl P.; Rabalski I. Composition and antioxidant properties of anthocyanin-rich products prepared from purple wheat. Food Chem, 2018, 254, 13-19.
- Abdel-aal E.-S.; Hucl P.; Shipp J.; Rabalski I. Compositional differences in anthocyanins from blue- and purple- grained spring wheat grown jn four environments in central Saskatchewan. Cereal Chemistry, 2016, 93, 32-38.
- Araus JL, Cairns JE. Field high-throughput phenotyping: the new crop breedingfrontier. Trends Plant Sci 2014;19(1):52–61.
- Arbuzova V.S.; Maystrenko O.I.; Popova O.M. Development of near-isogenic lines of the common wheat cultivar ‘Saratovskaya 29’. Cereal Res. Commun., 1998, 26, 39–46.
- Armstrong JM. Hybridization ofTriticumandAgropyron: I. Crossing results anddescription of the first generation hybrids. Can J Res 1936;14c(5):190–202.
- Asp T, Byrne S, Gundlach H, Bruggmann R, Mayer KFX, Andersen JR, et al.Comparative sequence analysis ofVRN1alleles ofLolium perennewith the co-linear regions in barley, wheat, and rice. Mol Genet Genomics 2011;286(5–6):433–7.
- Becker R, Wagoner P, Hanners GD, Saunders RM. Compositional, nutritionaland functional evaluation of intermediate wheatgrass (Thinopyrumintermedium). J Food Process Preserv 1991;15(1):63–77.
- Bell LW, Byrne F, Ewing MA, Wade LJ. A preliminary whole-farm economicanalysis of perennial wheat in an Australian dryland farming system. AgricSyst 2008;96(1–3):166–74.
- Bell LW, Wade LJ, Ewing MA. Perennial wheat: a review of environmental andagronomic prospects for development in Australia. Crop Pasture Sci 2010;61(9):679–90.
- Bohmdorfer S.; Oberlerchner J.T.; Fuchs C.; Rosenau T.; Graugsgruber H. Profiling and quantification of grain anthocyanins in purple pericarp x blue aleurone wheat crosses by high-performance thin-layer chromatography and densitometry. 2018, 14, 29.
- Borrill P, Connorton JM, Balk J, Miller AJ, Sanders D, Uauy C. Biofortification ofwheat grain with iron and zinc: integrating novel genomic resources andknowledge from model crops. Front Plant Sci 2014;5:53.
- Brand-Williams, W., Cuvelier, M.E. ve Berset, C. 1995. Use of a radical method to evaluate antioxidant activity. Lebensm. Wiss. u. Technology-Food Science and Technology, 28(1): 25-30.
- Buresova I.; Trojan V.; Helis M. Characteristics of flour and dough from purple and blue wheat grain. Slovak journal of food sciences, 2019, 13, 163-166.
- Bustos D.V.; Riegel R.; Calderini D.F. Anthocyanin content of grain in purple wheat is affected by grain position, assimilate availability and agronomic management. J. Cereal Science 2011, 55, 257–264.
- Cao S, Xu H, Li Z, Wang X, Wang D, Zhang A, et al. Identification andcharacterization of a novelAg. intermediumHMW-GS gene fromT. aestivum-Ag. intermediumaddition lines TAI-I series. J Cereal Sci 2007;45(3):293–301.
- Cattani DJ. Selection of a perennial grain for seed productivity across years:intermediate wheatgrass as a test species. Can J Plant Sci 2016;97(3):516–24.
- Chandi. G.K., & Seetharaman, K. (2012). Optimization of gluten peak tester: A statistical approach. Journal of Food Quality, 35, 69-75.
- Chen Q, Conner RL, Li HJ, Graf R, Laroche A, Li YH, et al. Genomiccharacterization of new sources of resistance to both wheat streak mosaicvirus and wheat curl mite in wheat–Thinopyrumpartial amphiploids. J GenetBreed 2003;57:155–64.
- Chen Q. Detection of alien chromatin introgression fromThinopyrumintowheat using S genomic DNA as a probe—a landmark approach forThinopyrumgenome research. Cytogenet Genome Res 2005;109(1–3):350–9.
- Colmer TD, Munns R, Flowers TJ. Improving salt tolerance of wheat and barley:future prospects. Aust J Exp Agric 2006;45(11):1425–43.
- Cooney D, Kim H, Quinn L, Lee MS, Guo J, Chen SL, et al. Switchgrass as abioenergy crop in the Loess Plateau, China: potential lignocellulosic feedstockproduction and environmental conservation. J Integr Agric 2017;16(6):1211–26.
- Cox S, Nabukalu P, Paterson AH, Kong WQ, Nakasagga S. Development ofperennial grain sorghum. Sustainability 2018;10(1):172.
- Cox TS, Bender M, Picone C, Van Tassel DL, Holland JB, Brummer EC, et al.Breeding perennial grain crops. Crit Rev Plant Sci 2002;21(2):59–91.
- Cox TS, Van Tassel DL, Cox CM, DeHaan LR. Progress in breeding perennialgrains. Crop Pasture Sci 2010;61(7):513–21.
- Culman SW, Snapp SS, Ollenburger M, Basso B, DeHeen LR. Soil and waterquality rapidly responds to the perennial grain Kernza wheatgrass. Agron J2013;105(3):735–44.L. Cui et al./Engineering 4 (2018) 507–513511
- Curwen-Mcadams C, Jones SS. Breeding perennial grain crops based on wheat.Crop Sci 2017;57(3):1172–88.
- Daotong Lia, Pengpu Wanga, Yinghua Luob, Mengyao Zhaoa, and Fang Chena 2017 Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. CRITICAL REVIEWS IN FOOD SCIENCE AND NUTRITION 2017, VOL. 57, NO. 8, 1729–1741.
- Davies CL, Waugh DL, Lefroy EC. Variation in seed yield and its components inthe Australian native grassMicrolaena stipoidesas a guide to its potential as aperennial grain crop. Aust J Agric Res 2005;56(3):309–16.
- DeHaan LR, Van Tassel DL, Anderson JA, Asselin SR, Barnes R, Baute GJ, et al.A pipeline strategy for grain crop domestication. Crop Sci 2016;56(3):917–30.
- DeHaan LR, Van Tassel DL. Useful insights from evolutionary biology fordeveloping perennial grain crops. Am J Bot 2014;101(10):1801–19.
- DeHaan LR, Wang SW, Larson SR, Cattani DJ, Zhang XF, Kantarski T. Currentefforts to develop perennial wheat and domesticateThinopyrum intermediumas a perennial grain. In: Batello C, Wade L, Cox S, Pogna N, Bozzini A, ChoptianyJ, editors. Perennial crops for food security. Proceedings of the FAO ExpertWorkshop. Rome: FAO; 2014. p. 72–89.
- Department of Economic and Social Affairs of the United Nation. The 2017revision of word population prospect. Report. New York: United Nations; 2017.Report No.: ESA/P/WP/248.
- Dong YS, Zhou RH, Xu SJ, Li LH, Cauderon Y, Wang RRC. Desirablecharacteristics in perennial Triticeae collected in China for wheatimprovement. Hereditas 1992;116(1–2):175–8.
- Erayman M.; Sandhu D.; Sidhu D.; Dilbirligi M.; Baenziger P.S.; Gill K.S. Demarcating the gene-rich regions of the wheat genome. Nucl. Acids Res. 2004, 32, 3546–3565.
- Eswaran H, Beinroth F, Reich P. Global land resources and population-supporting capacity. Am J Altern Agric 1999;14(3):129–36.
- Fiocco D.B.M.; De Simone V.; De Leonardis A.M.; Giovaniello V.; Del Nobile M.A.; Padalino L.; Lecce L.; Borelli G.M.; De Vita P. Use of purple durum wheat to produce naturally functional fresh and dry pasta. Food Chem 2016, 205, 187-195.
- Fradkin M, Ferrari MR, Ferreira V, Grassi EM, Greizerstein EJ, Poggio L.Chromosome and genome composition of aTriticumThinopyrum hybrid byclassical and molecular cytogenetic techniques. Genet Resour Crop Evol2012;59(2):231–7.
- Gantzer CJ, Anderson SH, Thompson AL, Brown JR. Estimating soil erosion after100 years of cropping on Sanborn Field. J Soil Water Conserv 1990;45(6):641–4.
- Gao, L., Wang, S., Oomah, B.D. and Mazza, G.2002. Wheat Quality: Antioxidant Activity of Wheat Millstreams, in:Wheat Quality Education. P. Ng and C.W. Wrigley, eds. AACC International, 219-233, St Paul, MN.
- Gazza L, Galassi E, Ciccoritti R, Cacciatori P, Pogna NE. Qualitative traits ofperennial wheat lines derived from differentThinopyrumspecies. GenetResour Crop Evol 2016;63(2):209–19.
- Gelfand I, Sahajpal R, Zhang X, Izaurralde RC, Gross KL, Robertson GP.Sustainable bioenergy production from marginal lands in the US Midwest.Nature 2013;493(7433):514–7.
- Glover JD, Reganold JP, Bell LW, Borevitz J, Brummer EC, Buckler ES, et al.Increased food and ecosystem security via perennial grains. Science 2010;328(5986):1638–9.
- Gordeeva E.I.; Shoeva O.Yu.; Yudina R.S.; Kukoeva T.V.; Khlestkina E.K. Effect of Seed Pre-sowing Gamma-irradiation Treatment in Bread Wheat Lines Differing by Anthocyanin Pigmentation. Cereal Res Commun., 2018, 46, 41-53.
- Gordeeva E.I.; Shoyeva O.Y.; Khlestkina E.K. Marker-assisted development of bread wheat near-isogenic lines carrying various combinations of purple pericarp (Pp) alleles. Euphytica, 2015, 203, 469-476.
- Grausgruber H.; Atzgersdorfer K.; Bohmdorefr S. Purple and blue wheat – health-promoting grains with increased antioxidant activity. Cereal food world, 2018, 63, 217-220.
- Harmoney KR. Cool-season grass biomass in the southern mixed-grass prairieregion of the USA. BioEnergy Res 2015;8(1):203–10.
- Hayes RC, Newell MT, DeHaan LR, Murphy KM, Crane S, Norton MR, et al.Perennial cereal crops: an initial evaluation of wheat derivatives. Field CropsRes 2012;133:68–89.
- Henderson J. W.,Ricker R. D., Bidlingmeyer B. A., ve Woodward C. 2000. Rapid, accurate, sensitive, and reproducible HPLC analysis of amino acids. https://www.agilent.com/cs/library/chromatograms/59801193.pdf erişim: 01.10.2017.
- Hong M.J.; Kim D.Y.; Nam B.M.; Ahn J.-W.; Kwon S.-J.; Seo Y.W.; Kim J.-B.; Characterization of novel mutants of hexaploid wheat (Triticum aestivum) with various depths of purple grain color and antioxidant capacity. J Sci Food Agric. 2019, 99, 55-63.
- Hosseinian F.S.; Li W.; Beta T. Measurement of anthocyanins and other phytochemicals in purple wheat. Food Chem. 2008, 109, 916-924.
- Hrivna L.; Zigmundova V.; Buresova I.; Maco R.; Vyhnanek T.; Trojan V. Pheological properties of dough and baking quality of products using coloured wheat. Plant Soil Envion. 2018, 64, 203-208.
- Jauhar PP. Multidisciplinary approach to genome analysis in the diploidspecies,Thinopyrum bessarabicumandTh. elongatum(Lophopyrum elongatum),of the Triticeae. Theor Appl Genet 1990;80(4):523–36.
- Jia JZ, Li HJ, Zhang XY, Li ZC, Qiu LJ. Genomics-based plant germplasm research(GPGR). Crop J 2017;5(2):166–74.
- Jones JM, Engleson J. Whole grains: benefits and challenges. Annu Rev Food SciTechnol 2010;1:19–40.
- Jones TA, Zhang XY, Wang RRC. Genome characterization of MT-2 perennialand OK-906 annual wheatintermediate wheatgrass hybrids. Crop Sci1999;39(4):1041–3.
- Jungers JM, DeHaan LR, Betts KJ, Sheaffer CC, Wyse DL. Intermediatewheatgrass grain and forage yield responses to nitrogen fertilization. Agron J2017;109(2):462–72.
- Kaçar, B., and Inal. A., 2008. Bitki Analizleri. Nobel. 912p. ISBN:978-605-395-036-3.
- Kantar MB, Tyl CE, Dorn KM, Zhang X, Jungers JM, Kaser JM, et al. Perennialgrain and oilseed crops. Annu Rev Plant Biol 2016;67:703–29.
- Kantarski T, Larson S, Zhang X, DeHaan L, Borevitz J, Anderson J, et al.Development of the first consensus genetic map of intermediate wheatgrass(Thinopyrum intermedium) using genotyping-by-sequencing. Theor Appl Genet2017;130(1):137–50.
- Kasem S, Waters DL, Rice N, Shapter FM, Henry RJ. Whole grain morphology orAustralian rice species. Plant Genet Resour 2010;8(1):74–81.
- Kassegn H.H. Determination of proximate composition and bioactive compinds of the Abyssinian purple wheat. Cogent food & agriculture, 4, 1421415.
- Khlestkina E.K.; Gordeeva E.I.; Arbuzova V.S. Molecular and functional characterization of wheat near-isogenic line ‘i:S29Ra’ having intensive anthocyanin pigmentation of the coleoptile, culm, leaves and auricles. Plant Breed., 2014, 133, 454-458
- Khlestkina E.K.; Pestsova E.G.; Röder M.S.; Börner A. Molecular mapping, phenotypic expression and geographical distribution of genes determining anthocyanin pigmentation of coleoptiles in wheat (Triticum aestivum L.). Theor. Appl. Genet., 2002, 104, 632-637.
- Khlestkina E.K.; Pshenichnikova T.A.; Röder M.S.; Börner A. Clustering anthocyanin pigmentation genes in wheat group 7 chromosomes. Cereal Res Commun, 2009, 37, 391-398.
- Khlestkina E.К.; Usenko N.I.; Gordeeva E.I.; Stabrovskaya O.I.; Sharfunova I.B.; Otmakhova Y.S. Evaluation of wheat products with high flavonoid content: justification of importance of marker-assisted development and production of flavonoid-rich wheat cultivars. Vavilovskii Zhurnal Genetiki i Selektsii, 2017, 21, 545-553. DOI 10.18699/VJ17.25-o
- Knievel D.C.; Abdel-Aal E.-S.; Rabalski I.; Nakamura T.; Hucl P. Grain color development and the inheritance of high anthocyanin blue aleurone and purple pericarp in spring wheat. Journal of Cereal Science 2009, 50, 113-120.
- Kryger S.; Morlock G.E. Fingeprinting and characterization of anthocyanins in 94 colored wheat varieties and blue aleurone and purple pericarp wheat crosses. J of Chromatography A, 2018, 1538, 75-85.
- Lam HM, Remais J, Fung MC, Xu L, Sun SSM. Food supply and food safety issuesin China. Lancet 2013;381(9882):2044–53.
- Lammer D, Cai X, Arterburn M, Chatelain J, Murray T, Jones S. A singlechromosome addition fromThinopyrum elongatumconfers a polycarpic,perennial habit to annual wheat. J Exp Bot 2004;55(403):1715–20.
- Larkin PJ, Newell MT, Hayes RC, Aktar J, Norton MR, Moroni SJ, et al. Progress indeveloping perennial wheats for grain and grazing. Crop Pasture Sci 2014;65(11):1147–64.
- Larkin PJ, Newell MT. Perennial wheat breeding: current germplasm and a wayforward for breeding and global cooperation. In: Batello C, Wade L, Cox S,Pogna N, Bozzini A, Choptiany J, editors. Perennial crops for food security.Proceedings of the FAO Expert Workshop. Rome: FAO; 2014. p. 39–53.
- Lenser T, Theißen G. Molecular mechanisms involved in convergent cropdomestication. Trends Plant Sci 2013;18(12):704–14.
- Li H, Wang X.Thinopyrum ponticumandTh. intermedium: the promising sourceof resistance to fungal and viral diseases of wheat. J Genet Genomics 2009;36(9):557–65.
- Li HJ, Conner RL, Murray TD. Resistance to soil-borne diseases of wheat:contributions from the wheatgrassesThinopyrum intermediumandTh.ponticum. Can J Plant Sci 2008;88(1):195–205.
- Li HJ, Cui L, Li HL, Wang XM, Murray TD, Conner RL, et al. Effective resources inwheat and wheat-derivatives for resistance toHeterodera filipjeviin China.Crop Sci 2012;52(3):1209–17.
- Li Q, Li L, Yang X, Warburton ML, Bai G, Dai J, et al. Relationship, evolutionaryfate and function of two maize co-orthologs of riceGW2associated with kernelsize and weight. BMC Plant Biol 2010;10:143.
- Li W.; Beta T. Flour and bread from black-, purple-, and blue-colored wheats. Flour and breads and their fortification in health and disease prevention. 2011. Academic press. 59-67.
- Li W.; Pickard M.D.; Beta T. Evaluation of antioxidant activity and electronic taste and aroma properties of antho-beers from purple grain. J Agric Food Chem 2007, 55, 8958-8966.
- Li X.; Lv X.; Wang X.; Zhang M.; Ren M. Effects of abiotic stress on anthocyanin accumulation and grain weight in purple wheat. Crop and pasture, 2018, 69, 1208-1214.
- Li YX, Zhang WF, Ma L, Wu L, Shen JB, Davies WJ, et al. An analysis of China’sgrain production: looking back and looking forward. Food Energy Secur 2014;3(1):19–32.
- Li Z, Li B, Tong Y. The contribution of distant hybridization with decaploidAgropyron elongatumto wheat improvement in China. J Genet Genomics2008;35(8):451–6.
- Lou H, Dong L, Zhang K, Wang DW, Zhao M, Li Y, et al. High-throughput miningof E-genome-specific SNPs for characterizingThinopyrum elongatumintrogressions in common wheat. Mol Ecol Resour 2017;17(6):1318–29.
- Ma D.; Zhang J.; Hou J.; Li Y.; Huang X.; Wang C.; Lu H.; Zhu Y.; Guo T. Evaluation of yield, processing quality and nutritional quality in different-colored wheat grains under nitrogen and phosphorus application. Crop Science, 2018, 58, 402-415.
- Ma D.; Zhang J.; Li Y.; Wang C. Quality of noodles made from colour-grained wheat. Czech J Food Sci, 2018, 36, 314-320.
- Ma XF, Gustafson JP. Allopolyploidization-accommodated genomic sequencechanges in triticale. Ann Bot 2008;101(6):825–32.512L. Cui et al./Engineering 4 (2018) 507–513
- Marti A, Bock JE, Pagani MA, Ismail B, Seetharaman K. Structuralcharacterization of proteins in wheat flour doughs enriched withintermediate wheatgrass (Thinopyrum intermedium) flour. Food Chem2016;194:994–1002.
- Marti A, Qiu X, Schoenfuss TC, Seetharaman K. Characteristics of perennialwheatgrass (Thinopyrum intermedium) and refined wheat flour blends: impacton rheological properties. Cereal Chem 2015;92(5):434–40.
- Martinek P.; Jirsa O.; Vaculova K.; Chrpova J.; Watanabe N.; Buresova V.; Kopecky D.; Stiasna K.; Vyhnanek T.; Trojan V. Use of wheat gene resources with different colour in breeding. 64 Tagung der Vereiningung der pflanzenzuchter und saatgutkaufleute Osterreichs, 2013, 1-4
- Meng QF, Hou P, Wu L, Chen XP, Cui ZL, Zhang FS. Understanding productionpotentials and yield gaps in intensive maize production in China. Field CropsRes 2013;143:91–7.
- Monfreda C, Ramankutty N, Foley JA. Farming the planet: 2. Geographicdistribution of crop areas, yields, physiological types, and net primaryproduction in the year 2000. Global Biogeochem Cycles 2008;22(1):1–19.
- Murphy KM, Hoagland LA, Reeves PG, Baik BK, Jones SS. Nutritional and qualitycharacteristics expressed in 31 perennial wheat breeding lines. Renew AgricFood Syst 2009;24(4):285–92.
- Newell MT, Hayes RC. An initial investigation of forage production and feedquality of perennial wheat derivatives. Crop Pasture Sci 2017;68(12):1141–8.
- Nkonya E, Mirzabaev A, von Braun J. Economics of land degradation andimprovement: an introduction and overview. In: Nkonya E, Mirzabaev A,vonBraun J, editors. Economics of land degradation and improvement—a globalassessment for sustainable development. Berlin: Springer; 2016. p. 1–14.
- Pask, AJD., Pietragalla, J., Mullan, DM. and Reynolds, MP. (Eds.) (2012) Physiological Breeding II: A Field Guide to Wheat Phenotyping. Mexico, D.F.: CIMMYT.
- Pasqualone A.; Bianco A.M.; Paradiso V.M.; Summo C.; Gabarcorta G.; Caponio F.; Blanco A. Production and characterization of functional biscuits obtained from purple wheat. Food Chem 2015, 180, 64-70.
- Paznocht L.; Kotilkova Z.; Orsak M.; Lachman J.; Martinek P. Carotenoid changes og colored-grain wheat flours during bun-making. Food Chem. 2019, 277, 727-734.
- Peto FH. Hybridization ofTriticumandAgropyron: II. Cytology of the maleparents and F1generation. Can J Res 1936;14c(5):203–14.
- Pimentel D, Cerasale D, Stanley RC, Perlman R, Newman EM, Brent LC, et al.Annual vs. perennial grain production. Agric Ecosyst Environ 2012;161:1–9.
- Randall GW, Mulla DJ. Nitrate nitrogen in surface waters as influenced byclimatic conditions and agricultural practices. J Environ Qual 2001;30(2):337–44.
- Robinson MD, Murray TD. Genetic variation ofwheat streak mosaic virusin theUnited States Pacific Northwest. Phytopathology 2013;103(1):98–104.
- Röder M.S., Huang X.Q., Börner A. Fine mapping of the region on wheat chromosome 7D controlling grain weight // Funct. Integr. Genomics. 2008. V. 8. P. 79-86.
- Sanderson MA, Adler PR. Perennial forages as second generation bioenergycrops. Int J Mol Sci 2008;9(5):768–88.
- Scheinost PL, Lammer DL, Cai XW, Murray TD, Jones SS. Perennial wheat: thedevelopment of a sustainable cropping system for the US Pacific Northwest.Am J Altern Agric 2001;16(4):147–51.
- Schulz-Schaeffer J, Haller SE. Registration of montana-2 perennialAgrotriticum intermediodurumKhizhnyak. Crop Sci 1987;27(4):822–3.
- Shao Y, Xu F, Sun X, Bao J, Beta T. Phenolic acids, anthocyanins, and antioxidant capacity in rice (Oryza sativa L.) grains at four stages of development after flowering. Food chemistry. 2014 Jan 15;143:90-6.
- Shapter FM, Cross M, Ablett G, Malory S, Chivers IH, King GJ, et al. High-throughput sequencing and mutagenesis to accelerate the domestication ofMicrolaena stipoidesas a new food crop. PLoS One 2013;8(12):e82641.
- Shoeva O.Y.; Gordeeva E.I.; Khlestkina E.K. The Regulation of Anthocyanin Synthesis in the Wheat Pericarp. Molecules. 2014, 19, 20266-20279
- Shoeva O.Yu.; Gordeeva E.I.; Arbuzova V.S.; Khlestkina E.K. Anthocyanins Participate in Protection of Wheat Seedlings from Osmotic Stress. Cereal Res Comm, 2017, 45, 47-56.
- Shoeva O.Yu.; Khlestkina E.K. Anthocyanins participate in the protection of wheat seedlings against cadmium stress. Cereal Res Commun., 2018, 46, 242-252.
- Singleton, V.L. and Rossi, J.A. 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungistic acid reagent. American Society for Enology and Viticulture,16,144-158.
- Smith DC. Intergenetic hybridization ofTriticumand other grasses, principallyAgropyron. J Hered 1943;34(7):219–24.
- Su Z, Hao C, Wang L, Dong Y, Zhang X. Identification and development of afunctional marker ofTaGW2associated with grain weight in bread wheat(Triticum aestivumL.). Theor Appl Genet 2011;122(1):211–23.
- Sun SC. Pursuit and exploration. Beijing: China Agriculture Press; 2015.Chinese.
- Sun SC. The approach and methods of breeding new varieties and new speciesfromAgrotriticumhybrids. Acta Agron Sin 1981;7(1):51–7. Chinese.
- Sun Y, Sun SC, Liu SX, Yan GY, Guo Q. Study on varieties breeding and selectionof perennial wheat. Seed 2011;30(4):21–6. Chinese.
- Suneson CA, Sharkawy AE, Hall WE. Progress in 25 years of perennial wheatdevelopment. Crop Sci 1963;3(5):437–9.
- Syed Jafar S.; Baron J.; Siebenhandl-Ehn S.; Rosenau T.; Bohmdorfer S.; Graugsgruber H. Increased anthocyanin content in purple pericarp x blue aleurone wheat crosses. Plant Breeding, 2013, 132, 546-552.
- Sykes VR, Allen FL, DeSantis AC, Saxton AM, Bhandari HS, West DR, et al.Efficiency of spaced-plant selection in improving sward biomass and ethanolyield in switchgrass. Crop Sci 2017;57(1):253–63.
- Sytar O.; Bosko P.; Zivcak M.; Brestic M.; Smetanska I. Bioactive phytochemicals and antioxidant properties of the grain and sprouts of colored wheat genotypes. Molecules. 2018, 23, 2282.
- Tereshchenko O.Y,; Gordeeva E.I.; Arbuzova V.S.; Börner A.; Khlestkina E.K. The D genome carries a gene determining purple grain colour in wheat. Cereal Res Commun, 2018, 40, 334–341
- Tereshchenko O.Y.; Arbuzova V.S.; Khlestkina E.K. Allelic state of the genes conferring purple pigmentation of different wheat organs predetermines tramscriptional activity of the anthocyanin biosynthesis structural genes. J Cereal Sci 2013, 57, 10-13.
- Tian S-Q.; Chen Z.; Wei Y. Measurement of colour-grained wheat nutrient compunds and the application of combination technology in dough. J Cereal Sci, 2018, 63-67.
- Tsitsin NV. Remote hybridization as a method of creating new species andvarieties of plants. Euphytica 1965;14(3):326–30.
- Usenko N.I.; Khlestkina E.К.; Asavasanti S.; Gordeeva E.I.; Yudina R.S.; Otmakhova Y.S. Possibilities of Enriching Food Products with Anthocyanins by Using New Forms of Cereals. Foods and Raw Materials, 2018, 6, 128–135. DOI: 10.21603/2308-4057-2018-1-128-135.
- Wagoner P, Schaeffer JR. Perennial grain development: past efforts andpotential for the future. Crit Rev Plant Sci 1990;9(5):381–408.
- Wagoner P. Perennial grain new use for intermediate wheatgrass. J Soil WaterConserv 1990;45(1):81–2.
- Wang RRC, Larson SR, Jensen KB. Differential transferability of EST-SSRprimers developed from the diploid speciesPseudoroegneria spicata,Thinopyrum bessarabicum, andThinopyrum elongatum. Genome 2017;60(6):530–6.
- Watson A, Ghosh S, Williams MJ, Cuddy WS, Simmonds J, Rey MD, et al. Speedbreeding is a powerful tool to accelerate crop research and breeding. NatPlants 2018;4:23–9.
- Weik L, Kaul HP, Kübler E, Aufhammer W. Grain yields of perennial grain cropsin pure and mixed stands. J Agron Crop Sci 2002;188(5):342–9.
- Xu YB, Crouch JH. Marker-assisted selection in plant breeding: frompublications to practice. Crop Sci 2008;48(2):391–407.
- Zeven A.C. 1991 Wheats with purple and blue grain. Euphytica, 56; 243-258.
- Zhang SL, Hu J, Yang CD, Liu HT, Yang F, Zhou JH, et al. Genotype byenvironment interactions for grain yield of perennial rice derivatives (OryzasativaL./Oryza longistaminata) in southern China and Laos. Field Crops Res2017;207:62–70.
- Zhang SL, Wang WS, Zhang J, Ting Z, Huang WQ, Xu P. The progression ofperennial rice breeding and genetics research in China. In: Batello C, Wade L,Cox S, Pogna N, Bozzini A, Choptiany J, editors. Perennial crops for foodsecurity. Proceedings of the FAO Expert Workshop. Rome: FAO; 2014. p.27–38.
- Zhang X, Dong Y, Wang RRC. Characterization of genomes and chromosomes inpartial amphiploids of the hybridTriticum aestivumThinopyrum ponticumbyin situhybridization, isozyme analysis, and RAPD. Genome 1996;39(6):1062–71.
- Zhang X, Sallam A, Gao L, Kantarski T, Poland J, DeHaan LR, et al. Establishmentand optimization of genomic selection to accelerate the domestication andimprovement of intermediate wheatgrass. Plant Genome 2016;9(1):1–18.
- Zhang XF, DeHaan LR, Higgins L, Markowski TW, Wyse DL, Anderson JA. Newinsights into high-molecular-weight glutenin subunits and sub-genomes ofthe perennial cropThinopyrum intermedium(Triticeae). J Cereal Sci 2014;59(2):203–10.
- Zhao HB, Zhang YM, Shi CL, Yan XD, Tian C, Li YP, et al. Development andcytogenetic analysis of perennial wheat in cold region. Acta Agron Sin 2012;38(8):1378–86. Chinese.
- Zhao XQ, Zhang T, Huang LY, Wu HM, Hu FY, Zhang F, et al. Comparativemetabolite profiling and hormone analysis of perennial and annual rice. J PlantBiol 2012;55(1):73–80.
- Zhu KX, Lian CX, Guo XN, Peng W, Zhou HM. Antioxidant activities and total phenolic contents of various extracts from defatted wheat germ. Food Chemistry. 2011 Jun 1;126(3):1122-6.
- Zrckova M.; Capuchova I.; Eliasova M.; Paznoch L.; Pasderu K.; Dvorak P.; Konvalina P.; Orsak M.; Sterba Z. The effect of genotypes, weather conditions and cropping system on antioxidant activity and content of selected antioxidant compunds in wheat with colored grain. Plant Spin Eviron. 2018, 64, 530-538.