
Титульный лист и исполнители
РЕФЕРАТ
Отчет 48 с., рис. 18, табл. 23, источн. 33.
СЕВЕРНЫЕ ДОМАШНИЕ ОЛЕНИ, ГЕНЕТИЧЕСКОЕ РАЗНООБРАЗИЕ, СЕЛЕКЦИОННО-ПЛЕМЕННАЯ РАБОТА, ПОДБОР, ПРОДУКТИВНЫЕ КАЧЕСТВА, ПОРОДЫ ОЛЕНЕЙ, БОНИТИРОВКА.
Объектом исследования являются домашние северные олени.
Цель работы – разработка базы данных биологических, продуктивных особенностей и генетической структуры пород домашних северных оленей Республики Саха (Якутия).
В процессе работы проводились сбор биологического материала, бонитировка и взвешивание эвенской, чукотской пород домашних северных оленей. По микросателлитным локусам ДНК генотипировано 178 оленей эвенской породы, 144 оленей чукотской породы, в том числе 54 завезенных с Чукотского АО.
Проведено взвешивание и взятие промеров у 498 домашних северных оленей, в том числе 106 оленей чукотской породы.
В результате экспериментальных работ получены данные по аллельному разнообразию 14 микросателлитных локусов: Rt6 , BMS1788, Rt 30, Rt1 , Rt9 , C143 , Rt7 , OheQ , FCB193 , C217 , Rt24 , C32 , BMS745 , NVHRT16 , T40 , C276. Установлены межпородные различия по количеству и по распределению частот встречаемости в группах животных различной породной принадлежности.
По результатам проведенных исследований разработана база данных биологических, продуктивных особенностей и генетической структуры пород домашних северных оленей Республики Саха (Якутия) (http://nauka.ysaa.ru/ckp/baza-dannih/).
ВВЕДЕНИЕ
Оленеводство в России охватывает огромную территорию Крайнего Севера от Скандинавии до Берингова пролива. На огромных просторах Севера, Сибири и Дальнего Востока, которые занимают около 30% всей территории России, выпасается более 1 млн. 800 тыс. голов домашних оленей и около 1 млн. диких северных оленей (сородичей домашних оленей). Всего же в настоящее время на территории Российской Федерации северным домашним оленеводством занимаются 4 республики, 3 края, 6 областей и 4 автономных округов, расположенных на более чем 4 млн. кв. км, а это 1/4 часть России (табл. 1).
Таблица 1 – Поголовье северных оленей в Российской Федерации, тыс. гол.
| Субъект Российской Федерации | 1990 | 2000 | 2015 | 2018 |
| Российская Федерация, в том числе: | 2260,6 | 1196,7 | 1606,15 | 1838,7 |
| Республика Коми | 123,6 | 110,1 | 85,79 | 93,7 |
| Ненецкий автономный округ | 186,3 | 122,1 | 177,48 | 188,6 |
| Архангельская область
(без автономного округа) |
4,4 | 2,9 | 1,78 | 1,8 |
| Мурманская область | 77,3 | 61,3 | 56,24 | 58,1 |
| Ханты-Мансийский автономный округ
(Югра) |
47,1 | 26,5 | 40,57 | н/д |
| Ямало-Ненецкий автономный округ | 490,5 | 504,7 | 733,48 | 788,6 |
| Республика Бурятия | 2,1 | 0,6 | 0,58 | н/д |
| Республика Тыва | 8,1 | 1,2 | 3,54 | н/д |
| Забайкальский край | 9,2 | 0,7 | 3,06 | н/д |
| Красноярский край | 107,4 | 46,2 | 116,32 | н/д |
| Иркутская область | 3,9 | 1,0 | 0,94 | н/д |
| Республика Саха (Якутия) | 361,5 | 156,2 | 156,01 | 154,6 |
| Камчатский край | 167,1 | 35,9 | 46,84 | 47,1 |
| Хабаровский край | 42,7 | 7,7 | 6,08 | н/д |
| Амурская область | 12,9 | 6,2 | 6,7 | н/д |
| Магаданская область | 121,5 | 17,7 | 14,43 | 10,3 |
| Сахалинская область | 4,0 | 3,3 | 0,1 | н/д |
| Чукотский автономный округ | 491,0 | 92,5 | 156,14 | 155,1 |
Северным домашним оленеводством в республике занимаются в 20 из 34 муниципальных районов. Ведение оленеводства в четырех зонах, приспособленность к суровым условиям и способность оленей выпасаться круглый год за счет естественных кормовых угодий, являются специфическими особенностями, отличающих якутское оленеводство от оленеводства других районов Крайнего Севера России.
Важнейшим элементом экосистем Севера и неотъемлемой частью жизни и культуры коренных народов Республики Саха (Якутия)является северный олень, Rangifer tarandus. Данный вид – единственный представитель рода Северные олени (Rangifer), относящегося к семейству оленевых подотряда жвачных, и один из немногих видов животных, у которых дикая форма сосуществует с домашней. Выведенный народной селекцией, разными северными народностями России, породный состав домашних северных оленей был обоснован и апробирован учеными Якутского НИИ сельского хозяйства и утвержден Министерством сельского хозяйства СССР Приказом №212 от 23.08.1985 года.
Из 4-х утвержденных пород (эвенская, эвенкийская, чукотская и ненецкая.) в Республике Саха (Якутия) разводятся 3 породы оленей: эвенская, эвенкийская и чукотская (харгин). Эти породы отличаются направлением продуктивности, экстерьерно-конституциональными особенностями и приспособленностью к конкретным природно-климатическим условиям. [1]
Таким образом, оценка генетических ресурсов оленеводства, постоянное наблюдение за популяционными изменениями является необходимым условием для своевременных и эффективных действий, направленных на предотвращение исчезновения пород северных домашних оленей. Мероприятия по предотвращению генетической эрозии и исчезновения популяций будут более эффективными, если четко определены факторы, способствующие их проявлению.
1 ОЛЕНЕВОДСТВО В РЕСПУБЛИКЕ САХА (ЯКУТИЯ)
1.1 Современное состояние северного оленеводства
Республика Саха (Якутия) является, наряду с Ямало-Ненецким и Чукотским автономными округами одним из самых крупных оленеводческих регионов России. Оленеводство исторически всегда отличалось здесь сложившейся традиционно высокой культурой разведения оленей. Здесь сохранилась система стадного содержания оленей, при которой оленеводы круглосуточно круглогодично выпасают оленей, и в поисках корма непрерывно в течение года кочуют вместе со стадом по пастбищам.
За годы всеобщей перестройки экономики оленеводство Республики Саха (Якутия), как и другие регионы России, пережило период глубокого кризиса, в результате чего произошло разгосударствление крупных оленеводческих совхозов, прекращение государственной поддержки отрасли. В этот период в Республике в результате реформирования сельскохозяйственного производства и разрушения организационной и технологической структуры, во многих улусах олени были переведены в собственность мелких коллективных и крестьянских (родовых) хозяйств. [2].
В период с 1990 по 1996 гг. в оленеводстве Якутии был допущен значительный сброс поголовья домашних оленей, сокращение поголовья составило 121,2 тысяч голов – 33%, произошло снижение всех производственных показателей отрасли.
Ситуация в оленеводстве республики начала меняться после принятия Правительством мер, направленных на стабилизацию положения в оленеводстве. Так, за период с 1995 по 2004 гг. было принято более 20 постановлений, направленных на сохранение традиционных отраслей, в частности, на улучшение условий жизни и труда оленеводов, на повышение их моральной и материальной поддержки. Реализация этих постановлений и принятых в республике на его основе организационно-хозяйственных мер дали положительные сдвиги. Начиная с 2003 года, был приостановлен более чем 20-летний спад поголовья оленей. К концу 2006 года поголовье домашних северных оленей достигло 169 тыс. голов. С января 2007 года в Якутии снят мораторий на убой оленей, и оленеводческие хозяйства с любым количеством оленей получили право реализации оленей по своему усмотрению. В 2008 году для реализации на мясном рынке в республике проведен забой оленей в количестве 10609 голов, с живой массой 848,7 тонн. [3].
Разведением северного домашнего оленя в республике занимаются в 20 из 34 муниципальных районов.
По состоянию развития оленеводства оленеводческие районы республики условно делятся на три группы.
В первую группу входят улусы с наиболее устойчивым оленеводством: Булунский, Нижнеколымский, Оймяконский улусы и Усть-Янский улусы. В этой группе улусов численность оленей находится в относительно стабильном уровне.
Во вторую группу входят Анабарский, Алданский, Оймяконский, Жиганский, Момский, Нерюнгринский, Нижнеколымский, Оленекский, Кобяйский, Томпонский и Эвено-Бытантайский улусы, где есть необходимость увеличения численности северных домашних оленей и на этой основе создавать новые рабочие места, повышать уровень жизни коренного населения, который во многом зависит от состояния оленеводства.
В третью группу входят Аллаиховский, Абыйский, Верхнеколымский, Верхоянский, Среднеколымский и Усть-Майский улусы. В данных улусах стоит задача сохранения оленеводства, как основы уклада жизни коренных малочисленных народов Севера.
Всего в отрасли оленеводства заняты 106 оленеводческих хозяйств. В оленеводческих бригадах работают – 1361 человек (в 2018 году – 1494 человек), в том числе оленеводов – 824 и чумработников – 283.
На 01 января 2020 года по предварительным данным управлений сельского хозяйства улусов численность оленей во всех категориях хозяйств составляет 152 127 голов или 103,8 % к началу 2019 года (146 585 голов), в том числе поголовье важенок и сыриц составило 70 202 голов.
Получено 41 721 тугутов. Деловой выход тугутов составляет 52,0 %, что лучше показателя прошлого года на 4,2 %. Лучшие показатели по ходу отела на данном этапе в хозяйствах Анабарского (72,4 %), Нижнеколымского (71,7 %) и Усть-Янского улусов (57,3 %).
Низкие показатели по деловому выходу тугутов отмечается в хозяйствах Верхнеколымского (39,9 %), Олекминского (40,4 %), Булунского (43,5 %), и Нерюнгринского (44,4 %). Данное положение связано с увеличением численности хищников и отмечается большой отход тугутов во время отела в следствие травежа медведями. В данных районах принимаются меры по защите от хищников.
Сохранность взрослого поголовья в среднем по республике составляет 80,9 % и выше показателя прошлого года на 8,3 %. Лучшие показатели по сохранности взрослого поголовья оленей по итогам 2019 года у Усть-Янского (89,2%), Нижнеколымского – 86,1 % и Момского улусов – 90,7%
Непроизводительный отход оленей составил 30 299 голов (2018 – 42429 гол) и составляет 20,7 %. В том числе падеж – 5396 (2018- 10517 гол), травеж – 11395 (2018 – 13825 гол) и потери – 11395 голов (2018 – 18087 гол). Непроизводительный отход оленей по сравнению с прошлым годом сокращен на 9,8 %.
1.2 Породный состав и породность оленей Якутии
В Республике Саха (Якутия) по данным Федеральной службы государственной статистики на 1 января 2020 года поголовье домашних северных оленей составляет 152,1 тыс. гол (рис. 1.).
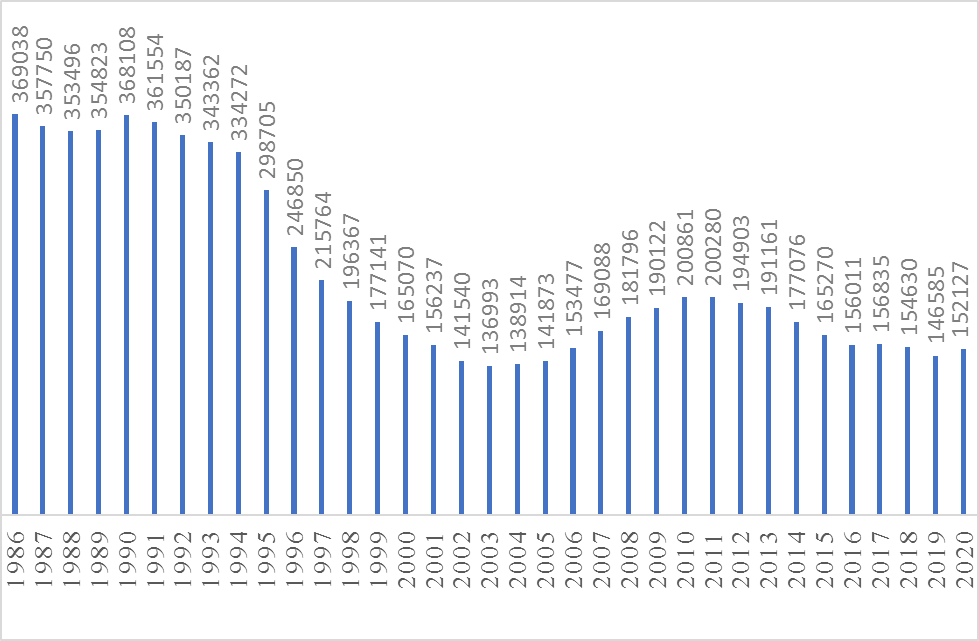
Рисунок 1 Динамика поголовья домашних северных оленей (гол.) в Республике Саха (Якутия) с 1986 по 2020 гг. на начало года
В Якутии из 4 утвержденных пород разводят 3 породы домашних северных оленей: эвенскую, эвенкийскую и чукотскую. Если две первые породы распространены по территории республики повсеместно, то чукотскую породу разводят только в Нижнеколымском районе. Доля эвенской породы – 60% (91,8 тыс.гол.), эвенкийской – 30% (45,1 тыс.гол.) и оленей чукотской породы в общей численности домашних оленей занимает только 10 % (15,1 тыс. гол.).
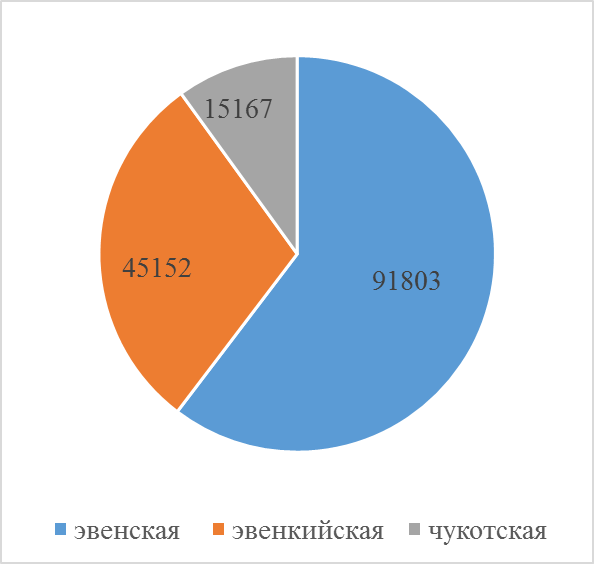
Рисунок 2 Поголовье оленей по породному составу, голов
Эвенская порода. Разводится в 11 улусах (районах) Республики Саха (Якутия) тундровой и горно-таежной зоны. Это самая многочисленная из существующих пород на территории республики, среди которых исследователи выделяют эвенских тундровых и таежных оленей. На начало 2020 года составляет 91803 голов.
Таблица 2 – Динамика поголовья эвенской породы в Якутии
| Улусы | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
| Всего по Республике Саха (Якутия) | 177076 | 165270 | 156011 | 156835 | 154630 | 146585 | 152127 |
| Абыйский | 626 | 703 | 446 | 515 | 20 | 59 | 79 |
| Булунский | 14153 | 13007 | 11526 | 12337 | 13351 | 14148 | 14366 |
| Верхнеколымс кий | 1203 | 1121 | 1016 | 1355 | 1386 | 1171 | 820 |
| Верхоянский | 4275 | 4350 | 3459 | 3273 | 2358 | 2469 | 2591 |
| Кобяйский | 9352 | 8553 | 7900 | 7211 | 7168 | 6371 | 6159 |
| Момский | 13904 | 14009 | 13186 | 10847 | 10872 | 10997 | 12761 |
| Оймяконский | 13237 | 9716 | 9333 | 10051 | 10270 | 7677 | 8303 |
| Среднеколымс кий | 2304 | 2172 | 2033 | 2080 | 1379 | 1371 | 1521 |
| Томпонский | 10354 | 10564 | 8405 | 8076 | 7543 | 6060 | 6412 |
| Усть-Янский | 20903 | 20117 | 21714 | 22774 | 22456 | 24203 | 25677 |
| Эвено- Бытантайский | 16182 | 14648 | 15371 | 15750 | 14477 | 13137 | 13114 |
| ИТОГО | 106493 | 98960 | 94389 | 94269 | 91280 | 87663 | 91803 |
| % от всего поголовья | 60 | 60 | 61 | 60 | 59 | 60 | 60 |
Имея общность происхождения, северные олени эвенской породы хорошо приспособлены к условиям содержания в тундровых и горно-таежных зонах Якутии.
Выпас оленей осуществляется крупными стадами, основное направление породы мясо-транспортная.
В отличие от других пород имеют разнообразную масть от белой, светло-серой до бурой и темно-бурой. Олени эвенской породы – животные преимущественно бурой масти с различными оттенками (80-90 %), встречаются темно-серые и светло-серые, менее распространены белые и пегие олени, редки с отметинами на морде, с белой головой и в «чулках». Все олени с хорошо выраженным типом породы, имеют легкое сухое сложение с плотной мускулатурой, крепкой конституцией, пропорциональным телосложением, без ярко выраженных мясных качеств, голова типичная для породы, рога хорошо развиты, половые признаки хорошо выражены, относятся к лептосомному типу.
Для экстерьера оленей эвенской породы характерно хорошее развитие туловища в длину и глубину при небольшом показателе ширины.
Индекс массивности 108,8-110,2%, глубокогрудности – 37,9-38,1%. Конечности средней длины, индекс длинноногости – 57,9-60,2%. Голова несколько длинная, но в лобной части не широкая.
Воспроизводительная способность животных удовлетворительная: яловость маток в оленеводческих хозяйствах республики составляет с среднем 15-16%, а в лучших хозяйствах и стадах, где проводится племенная работа, — 50-77%.
Мясные качества оленей эвенской породы характеризуется высокими показателями; при убое взрослых быков получают туши массой 53-62 кг, взрослых самок – 43-48 кг, убойный выход равняется соответственно 49,8 и 50,6%. Относительный выход массы внутренних органов составляет: сердца – 0,90-0,92%, легких – 1,38-1,43%, печени – 1,40-1,43%, почек – 0,22-0,23%.
В тушах телят 6-месячного возраста содержится мышечной ткани в среднем 68%, жировой – 4,10, соединительной – 7,84, костной – 19,62%; в тушах 2,5 летних животных соответственно 67,9%; 10,11%; 7,74%; 16,7%.
Консолидации недостатков в генотипе породы предшествовало в первую очередь возросшее транспортное использование оленей в прошлом для хозяйственных целей (почтовая и пассажирская связь, вывоз различных грузов). Формировалось множество транспортных стад оленей. Отбор животных вели в основном по рабочим качествам. Системы отбора оленей по мясным качествам не изучались, и им не придавали значения. И лишь в последние десятилетия А.Д. Мухачев впервые разработал научно обоснованную методику отбора оленей мясного (широкотелого) типа, позволяющую при том же поголовье увеличить выход мяса на 10-15%. [4].
Эвенкийская порода. Распространена на территории Алданского, Нерюнгринского, Олекминского, Оленекского, Жиганского, Усть-Майского и Горного улусов. В Анабарском улусе разводится тундровой тип эвенкийских оленей. Общая численность на начало года составляет 45152 голов.
Таблица 3 — Динамика поголовья эвенкийской породы в Якутии
| Улусы | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
| Всего по Республике Саха (Якутия) | 177076 | 165270 | 156011 | 156835 | 154630 | 146585 | 152127 |
| Алданский | 11444 | 9859 | 8123 | 7717 | 7878 | 7366 | 7428 |
| Анабарский | 13922 | 13825 | 14503 | 15407 | 16964 | 17682 | 18069 |
| Вилюйский | 187 | 214 | 216 | 179 | |||
| Горный | 280 | 304 | 225 | 230 | 253 | 57 | 48 |
| Жиганский | 5228 | 4004 | 3081 | 2709 | 2779 | 2931 | 2688 |
| Нерюнгри | 7106 | 6446 | 5797 | 5917 | 6047 | 6204 | 6110 |
| Олекминский | 4982 | 5412 | 5880 | 5503 | 5940 | 5911 | 5112 |
| Оленекский | 5463 | 4864 | 4187 | 4403 | 4591 | 5016 | 5287 |
| Усть-Майский | 550 | 632 | 543 | 567 | 571 | 649 | 410 |
| Итого | 49162 | 45560 | 42555 | 42632 | 45023 | 45816 | 45152 |
| % от всего поголовья | 28 | 28 | 27 | 27 | 29 | 31 | 30 |
Череп данной породы отличается наибольшей крупностью среди других пород. Исследователями А.Д.Мухачева, Ю.М.Макушева (1976) установлено, что наибольшая длина в среднем равна 406 мм, ширина 175 мм; у самок, соответственно – 362 мм и 160 мм. Для черепа характерна слабая выраженность перехода между мозговой и лицевой частями, неглубокое межглазничное вдавливание, мощное развитие затылочной области. [5]
Доминирующие масти светло-бурая и серая.
Относительная скорость роста у оленей наивысшая до 6-месячного возраста, после чего она снижается. В этом возрасте у оленей эвенкийской породы масса телят самцов от взрослых составляет 43,41%, телят самок – 57,18% (Мухачев, Макушев, Яковлев, 1975). Средняя живая масса взрослых оленей составляет – 142,0 кг, самок – 102 кг. [6].
Олени эвенкийской породы имеют большую живую массу по сравнению с другими породами. Высокая живая масса соответствует их телосложению – это высокие животные с хорошо развитым в длину туловищем, глубокой грудью, с хорошо развитой мускулатурой и костяком. Средняя величина промеров (см: ) самца – высота в холке в холке 115, глубина – 49, ширина груди – 28, обхват груди – 136, косая длина туловища – 121, обхват пясти – 14,1, ширина в маклоках – 27; соответственно, самки – 104; 44; 25; 125; 111; 11,9; 24.
Изучение мясной продуктивности оленей эвенкийской породы (Мухачев, Ким, 1972; Мухачев, Машуров, 1977) установило, что масса туши самцов 6-месячного возраста – 34,80 кг; взрослых самцов – 75,17 кг; убойный выход – 49,22%; 48,79%. У самок соответственно – 30,64 кг; 53,01 кг и 48,7%; 48,97%.
В тушах 6-месячных телят самцов содержится мышечной ткани – 66,88%; жира – 7,7%; сухожилий 8,2%; костной – 17,22%. У взрослых соответственно – 65,65%; 10,67%; 9,01%; 14,67%. Площадь мышечного глазка и толщина мускульного волокна у молодняка – 16,86 см2 и 34,34 см2, содержание влаги в мясе 71,23 %; сухого вещества – 28,77%; калорийность 1 кг мяса – 1821 ккал. У взрослых соответственно – 25 см2 ,35 см2; 61,85 %; 30%, 2053 ккал. [7].
Чукотская порода. Домашних северных оленей разводят на территории Чукотского АО, Камчатского АО (за исключением хозяйств, занимающихся с эвенской породой). На территории Якутии разводится только в Нижнеколымском улусе, поголовье на начало 2020 года 15167 голов.
Таблица 4 — Динамика поголовья чукотской породы в Якутии
| Улусы | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 |
| Всего по Республике Саха (Якутия) | 177076 | 165270 | 156011 | 156835 | 154630 | 146585 | 152127 |
| Нижнеколымский | 20377 | 20130 | 19067 | 19920 | 18317 | 13094 | 15167 |
| % от всего поголовья | 12 | 12 | 12 | 13 | 12 | 9 | 10 |
Научными исследованиями установлено, что чукотская порода самая мелкоплодная среди других пород оленей Якутии. Так, средняя живая масса при рождении у самцов 6,4, у самок – 5,4 кг.
Однако в первые месяцы постнатального периода чукотские телята по интенсивности роста заметно превосходят оленей других пород: максимальный среднесуточный прирост у них достигает 450-510 граммов.
К 6-месячному возрасту самцы достигают живой массы 60,9 кг, самки – 58,3 кг, в 1,5-летнем возрасте – 86,7 кг и 79 кг. Взрослые самцы имеют массу 110,3-125,5 кг, самки 88,8-92,8 кг. Эта порода имеет высокие нагульно — нажировочные качества: в условиях короткого вегетационного периода чукотские олени способны полностью восстанавливать зимние потери и уже в сентябре набрать высшую упитанность. Эта биологическая особенность хозяйственно полезного признака позволяет полнее использовать корма на пастбищах и не зависеть от запасов и доступности ягельников. Генетически закрепленные в породе, эта особенность при частых гололедах, не только позволяет выживать чукотским оленям, но и служит основой дальнейшего воспроизводства в условиях растущей делихенизации пастбищ. Не менее важной хозяйственно-полезной особенностью чукотских оленей являются ранние сроки наступления гона и отела, благодаря чему телята у важенок харгин до массового лета гнуса и наступления жары хорошо нагуливается и прибавляют в весе. [8].
Более ярко, чем у других пород, в телосложении чукотской породы выражены особенности мясного типа: хорошо развиты в ширину, при относительной низкорослости, передняя и задняя часть туловища, что придает ему бочкообразный вид. Они более коротконоги (Индекс длинноногости 57,3-58,6%), с крепким костяком (индекс костистости 12-13,6%). Голова не длинная, широкая, что объясняется лучшим развитием черепа в ширину как в мозговом, так и лицевом отделах.
Высокие мясные качества в период массового, осеннего забоя обеспечиваются у чукотских оленей высокой нажировкой летом и преимущественным развитием задней части туловища. От самцов в 6-месячном возрасте получают тушу массой 32,5 кг, убойный выход составляет 53,4%, от самок – соответственно 31,6 кг и 53,6%. В 1,5 летнем возрасте эти показатели у самцов составили 45,3 кг и 52,2%, у самок – 41,2 кг и 52,9%, у взрослых чукотских самцов – 58,8 кг и 53,3%, у самок – 49,1 кг и 52,6%. [9].
1.3 Микросателлитные маркеры и их использование в животноводстве
В настоящее время изучение домашних северных оленей с точки зрения генетики остается малоизученным. Если геном крупного рогатого скота секвенирован и определены особенности генов и их расположение в хромосомах (Sharifzadeh A., Doosti A., 2012; Долматова И.Ю. и др., 2012; Перчун А.В. и др., 2014), то геном северного домашнего оленя секвенирован лишь частично (NCBI), а поиск молекулярно-генетических маркеров хозяйственно-полезных признаков находится на начальном этапе. [10, 11, 12].
Одним из последних исследований в области поиска генетических маркеров является работы ФБГНУ «Всероссийский научно-исследовательский институт генетики и разведения сельскохозяйственных животных» в результате их исследования обнаружено четыре однонуклеатидных полиморфизма (SNP) в гене соматотропина, 2 из которых находятся в кодирующей части (Крутикова и др.2016.). [13].
В своих исследованиях Харзинова В.Р. и др. (2017) показали возможность использования на северных оленях ДНК чипа Bovine SNP50 BeadChip для крупного рогатого скота, который включает в себя 54609 однонуклеотидных замен. Результаты использования данного чипа выявили 512 полиморфных SNP, которые дают возможность в полной мере судить о генетической структуре пород домашних северных оленей. [14].
На сегодняшний день, в изучении генетической структуры северных оленей используются микросателлитные маркеры позволяющие выявить эволюционые процессы, протекающие в популяциях диких и домашних северных оленей (Митрофанова и др. 2015). [15].
Микросателлитные локусы являются высоко вариабельными областями ДНК, которые состоят из тандемно повторяющихся последовательностей длиной от одного до шести нуклеотидов. Число повторяющихся мотивов составляет от 10 до 30. Локусы микросателлитов нестабильны и характеризуются высокой скоростью мутационных изменений, приводящих к увеличению или уменьшению числа повторов, поэтому многие микросателлиты являются вариабельными по числу повторяющихся единиц, которое различается у разных особей. Нет двух особей, которые имели бы одинаковую комбинацию длин микросателлитных аллелей. Уникальность генетических профилей по микросателлитным последовательностям ДНК позволяет их использовать для идентификации особей и установления родственных взаимоотношений при проведении генетической экспертизы происхождения (Cervini M., 2006; Carolino I., et al., 2009; Stevanovic J et al. 2010; Karima F. Et al., 2011). [16, 17, 18, 19]
Микросателлитные маркеры ДНК могут служить для генетической идентификации пород, типов, линий сельскохозяйственных животных, определения генетической структуры и оценки генетических расстояний между группами животных, для оценки величины и направления генного потока между популяциями, для определения эффективного размера популяции для исчезающих и малочисленных пород сельскохозяйственных животных (Breen, M et al. 1997; Cervini M., 2006;Pevzner P.A., et al., 2006; Compeau Ph.E.C., et al.; Radko A. Rychlik T., 2009). [20, 21, 22, 23]
Особенности аллелофонда микросателлитных маркеров башкирской популяции симментальского скота исследовали И.Ю. Долматова и др. 2012, и результаты исследований по 12 локусам микросателлитов в сравнении с чистопородными и голштинизированной популяциями, показали, что башкирская популяция обладает относительно невысоким уровнем генетического разнообразия (7,08±0,72) (Долматова И.Ю и др. 2012). [11]
Результаты исследований генетической дифференциации внутрипородных линий уток по данным рестрикционного анализа митохондриальной ДНК, показали, что праймеры НМ13 и RM13 являются достаточно информативными для выявления межпородной и межлинейной дифференциации уток (Долматова И.Ю., 2006; Фисинин В.И. и др. 2006). [24, 25].
Полиморфизм молекулярно-генетических маркеров у овец романовской породы, с использованием в качестве праймеров последовательностей (GA)9C и (AG)9C, изучали Столповский Ю.А. и др. (2008). По результатам исследований пришли к выводу, что генотипирование животных с использованием этих молекулярно-генетических маркеров позволит контролировать и корректировать селекционную работу (Столповский и др., 2008). [26].
Также генетическую дифференциацию овец эдильбаевской породы с использованием праймеров (GA)9C, (AG)9C, (СТС)6С и (GAG)6С изучили Ельсукова и др. (2010). В ходе работы обнаружены достоверные отличия между внутрипородными типами бирликский и суюндукский по частоте встречаемости ISSR-PCR маркеров. Помимо этого, у овец этой породы обнаружены ранее не присутствовавшие, у других пород овец, фрагменты ДНК типичные для снежных баранов (Ельсукова и др. 2010). [27].
Оценка мультилокусной системы генетического анализа по ДНК-микросателлитам, выполненная на 11 группах свиней разных пород, показала высокую информативность (среднее число аллелей 5,15±0,33), этой системы. Система включает в себя 6 маркеров SW24, SW72, S0155, S0355, S0386 и SW951. Т.И. Тихомировой, предложена и доказана результативность этой системы при контроле достоверности происхождения (Проскурина Н.В., Тихомирова Т.И., 2007, Зиновьева Н.А. и др., 2008). [28, 29].
Также, Н.А. Зиновьева и др. (2009) изучив аспекты использования микросателлитов в свиноводстве, пришли к выводу, что микросателлитные профили можно использовать в популяционно-генетических исследованиях и при контроле достоверности происхождения (Зиновьева Н.И. и др., 2009; О.В. Харзинова, 2017). [30, 31].
2 РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
2.1. Материалы и методы исследований
Объект исследований. Объектом исследования послужили олени чукотской породы разводимые в Нижнеколымском районе СПК КРО «Турваургин» — 106 голов, в том числе закупленные с МП СХП БМР «Островное» — 54 головы (Чукотский АО). Олени эвенской породы – 392 головы, разводимые в племенном хозяйстве ФГУП «Ючюгейское» Оймяконского района.
Материалы исследования. Материалом для генетического анализа микросателлитных локусов послужили образцы ДНК, домашних северных оленей.
Методы исследования. Взрослых оленей взвешивали динамометром марки ДПУ-0,2-2. Экстерьерные особенности оленей определяли взятием промеров: а) мерной палкой — высота в холке, высота в локте, глубина груди, ширина груди за лопатками; б) мерной лентой — косая длина туловища, обхват груди, обхват пясти; в) мерным циркулем — длина головы, ширина в маклоках, косая длина зада. Все промеры брались с точностью 1 см (промер обхвата пясти — с точностью 0,1 см). Промеры брались у самцов перед гоном в августе, у самок в октябре. Пробонитировано 499 голов оленей. При обработке материала для характеристики пропорций телосложения использовался способ индексов.
Кровь для выделения ДНК отбирали из ярёмной вены в объёме 6 мл в вакуумные пробирки для гематологических исследований с ЭДТА К3. Выделение ДНК проводилось в учебной научно-исследовательской лаборатории ФГБОУ ВО Якутская ГСХА набором реагентов EXCELL BIOTECH (Разработка ГАУ «Технопарк Якутия», Россия).
Таблица 5 – Описание микросателлитных маркеров COrDIS RANGIFER
| Название маркера | Референсный аллель | Структура повтора |
| Rt6 | 19 | (СА)19 |
| BMS1788 | 17 | (АС)17 |
| Rt 30 | 15 | (АС)15 |
| Rt1 | 19 | (АС)19 |
| Rt9 | 21 | (АС)21 |
| FCB193 | 13 | (АС)13 |
| Rt7 | 11 | (АС)11 |
| BMS745 | 13 | (АС)13 |
| C 143 | 7 | (ATGG)7 |
| Rt24 | 21 | (АС)21 |
| OheQ | 20,3 | (TATC)17ATCTATCTATTTATC |
| C217 | 9 | CATC(CATG)5(CATC)3 |
| C32 | 14 | (ATCC)4 (ACCT)2 (ATCC)7 |
| NVHRT16 | 25 | (AC)5 AT (AC)4 ATGCGC (AC)12 |
| T40 | 13 | (ATCT)4 ACCT ATCT (ATCT)4 ACTG ACCT ATCT |
| C276 | 53 | (ТССА)5 ТССT TCCA TACG (TCCA)3 TCCT TCCA TCTG (TCCA)4 TCCG (ТССА)5 TCCT TCCA TCCG (ТССА)3 (TCCG)2 TGCA (TCCA)2TCCG TCCA |
Генотипирование проводили набором реагентов для мультиплексного анализа 16-ти микросателлитных маркеров северного оленя COrDIS Rangifer разработанная резидентом инновационного центра «Сколково» ООО «Гордиз» (таблица 5).
Статистическая обработка. При обработке экспериментальных данных использовали надстройку для Microsoft Excel — GeneAlex 6.51.
2.2. Характеристика эвенской породы
Формирование эвенской породы было положено эвенами, юкагирами, позднее якутами и коряками. Олень для них служил не только источником пищи, но широко использовали как транспортное животное, что наложило определенный отпечаток на его экстерьерно-конституциональные особенности. Эвенских оленей запрягают в нарты и ездят верхом. Эвенская порода оленей хорошо приспособлены к условиям содержания в лесотундровых, таежных и горно-таежных зонах Якутии. [32]
Олени эвенской породы занимают промежуточное положение между эвенкийской и чукотской породами. Они более приспособлены к горным районам, где летние пастбища расположены на высокогорьях, а зимние впадинах и долинах рек. Для них характерны выпас с короткими маршрутами кочевий с небольшим размером стад.
Основная масть светло-бурая. Телосложение более высокие с длинным и относительно узким туловищем, но с глубокой грудью, с длинным узким тазом, с длинной неширокой головой, с хорошо развитой мускулатурой и костяком.
Исследования проводили на 392 головы оленей эвенской породы в племенном хозяйстве ФГУП «Ючюгейское» Оймяконсого улуса (района) Республики Саха (Якутия).
Природно-климатические условия Оймяконского улуса Республики Саха (Якутия). Зоотехнические типы пастбищ северных оленей.
Муниципальное образование «Оймяконский улус (район)» находится на северо — востоке Республики Саха (Якутия) в пределах географических координат 64 с.ш. и 144 в.д. площадь – 92,2 тыс. кв. км., с восточной стороны граничит с Магаданской областью, с южной — Хабаровским краем, с западной стороны граничит с Томпонским улусом и с северной стороны- с Момским улусом. Через территорию улуса проходят горные массивы Черского и Верхоянского хребтов, на которых находится самая высокая точка в европейской части России- пик Муус — Хайа (высота 3011 метров).
Природные условия. Рельеф горный. На востоке улуса — Нерское плоскогорье; в центральной части — хребет Тас-Кыстабыт; на западе — Оймяконское нагорье, Эльгинское плоскогорье; на крайнем юго-западе — хребет Сунтар-Хаята; на севере — хребты горной системы Черского. Вся территория улуса находится в бассейне реки Индигирка.
Средняя годовая температура составляет по станции Оймякон – 16,6. а по станции Оймякон, аэропорт. -17,0.
Самая низкая температура воздуха по району Оймякона (-67.7) зафиксирована в Крест — Томторе 06.11 1933г. (ее отметил В.И.Попов):
В селе Оймякон находится полюс холода, где была зафиксирована одна из самых низких температур в северном полушарии −71,2 °C.
Средняя дата появления снежного покрова по станции Оймякон – 24.09, самая ранняя-24.08, самая поздняя 21.10, а по станции Оймякон, аэропорт, соответственно 6.10, 22.09, 21.10. Даты разрушения устойчивого снежного покрова: по станции Оймякон, аэропорт, соответственно 12, 30.04, 29.05
Даты схода снежного покрова: по станции Оймякон — средняя 11.04, самая ранняя 6.05, самая поздняя 31.05, а по станции Оймякон, аэропорт, соответственно 17.05. 04.05, 29.05.
Наибольшая высота снежного покрова по средним декадным величинам составляет: по станции Оймякон 29 см в марте, а по станции Оймякон, аэропорт 31 см в феврале — марте. Наибольшая за зиму высота снежного покрова составляет 43 см по станции Оймякон, аэропорт.
Число дней с грозой невелико и составляет: по станции Оймякон в среднем за год 10 (с мая по август) с максимумом в июне — июле, по станции Оймякон, аэропорт. -12 (с мая по сентябрь) с максимумом 5 в июле и 4 в июне. Наибольшее число дней с грозой в гол по станции Оймякон, аэропорт составляет 20 с максимумом (по 8 дней) в июне, июле, августе, что близко к числу дней, отмечающихся в Верхоянске и Якутске.
Вскрытие р. Индигирки в среднем происходит 21.05, самая ранняя дата 13.05 (1967г.) самая поздняя 3.06 (1947г.). Ледостав наступает в среднем 25.10 самый ранний срок — 7.10 (1941 г.) и самый поздний — 4.11 (1967г.). Средняя дата появления плавучего льда осенью 3.10 самая ранняя — 21.09 (1960 г.), а самая поздняя -13.10 (1961 г.).
Месячная сумма солнечной радиации на перпендикулярную поверхность имеет два максимума: 17,3 ккал/см2. Сумма радиации за год составляет 117,5 ккал/см 2. Максимум солнечной радиации на горизонтальную поверхность составляет 18.4 ккал/см2 в июне и 8,0-в апреле. Минимум 0,1 в январе и 0,0 в декабре, сумма за год 50,6 ккал/см2.
Основные зоотехнические типы оленьих пастбищ в Оймяконском улусе — различают следующие зоотехнические типы пастбищ: долины горных рек; равнины; крупнокустарниковые пастбища; лиственничное редколесье; лесные пастбища; пойменные и дерновые луга.
Оценка оленей по комплексу признаков экстерьера и конституции выявило: эвенская порода северных оленей по экстерьеру довольно высоконогие животные самцы в холке 112,1 см, самки 102,4 см, высота в локте составило у самцов 70,6, у самок 66,2, косая длина туловища у самцов 120,2 см, самки 109,7 см, обхват груди у самцов – 127,8 см, у самок – 110,8 см, обхват пясти у самцов 13,4 см, у самок – 12,2 см. (таблица 6,7).
Таблица 6 — Линейные промеры самцов (хоры) (n-31 гол)
| Показатели | средний | минимальный | максимальный |
| Высота в холке, см. | 112,1 | 112,1 | 112,0 |
| Косая длина туловища, см | 120,2 | 120,2 | 121,0 |
| Высота в локте, см | 70,6 | 70,6 | 70,0 |
| Обхват груди, см | 127,8 | 127,8 | 126,0 |
| Обхват пясти, см | 13,4 | 13,4 | 13,5 |
Отсутствие пороков в экстерьере и хорошее общее развитие эвенских оленей – это результат упорного труда эвенов, которые столетиями закрепляли за местными оленями необходимые хозяйственно-полезные признаки, хорошо приспособленных к условиям содержания в горно-таежной и лесотундровой зонах республики.
Таблица 7 – Линейные промеры самок (важенки) (n-362 гол)
| Показатели | средний | минимальный | максимальный |
| Высота в холке, см. | 102,4 | 96,0 | 105,0 |
| Косая длина туловища, см | 109,7 | 101,0 | 124,0 |
| Высота в локте, см | 66,2 | 60,0 | 76,0 |
| Обхват груди, см | 110,8 | 102,0 | 123,0 |
| Обхват пясти, см | 12,2 | 11,0 | 13,0 |
Таблица 8 – Индексы телосложения эвенской породы оленей
| Индексы, % | ФГУП «Ючюгейское» | По литературным данным | ||
| самцы | самки | самцы | самки | |
| Растянутости | 107,2 | 107,1 | 105,9 | 104,5 |
| Костистости | 11,9 | 11,9 | 11,9 | 11,5 |
| Массивности | 114,0 | 108,2 | 112,2 | 108,6 |
| Длинноногости | 62,9 | 64,6 | 58,1 | 59,2 |
| Сбитости | 106,3 | 101,0 | 106,0 | 103,9 |
По данным таблицы 7 индексы телосложения (%) у самцов: индекс растянутости 107,2; костистости 11,9; массивности 114,0; длинноногости 62,9; сбитости 106,3; у самок: растянутости 107,1; костистости 11,9; массивности 108,2; длинноногости 64,6; сбитости 101,0; (таблица 8).
Таблица 9 – Живая масса самцов эвенской породы оленей, кг
| Показатели | по литературным данным | ФГУП «Ючюгейское»
(n-31 гол) |
||
| средний | min | max | ||
| Живая масса, кг | 120,0 | 121,8 | 118,0 | 126,0 |
Средние показатели живой массы оленей эвенской породы ФГУП Ючюгейское» составили у самцов – 121,8 кг, у самок – 121,8 кг, которые коррелируют с литературными данными. (таблица 8, 9).
Таблица 10 – Живая масса самок эвенской породы оленей, кг
| Показатели | по литературным данным | ФГУП «Ючюгейское»
(n-362 гол) |
|||
| средний | min | max | |||
| Живая масса, кг | 92,0 | 94,3 | 85,0 | 108,0 | |
2.3. Характеристика чукотской породы
Большой труд в формировании чукотской породы вложили юкагиры, а позднее чукчи и коряки. В течение длительного времени олени чукотской породы совершенствовались по направлению мясной продуктивности. Выпас оленей проводили пешком, с целью облегчения окарауливания из стада постоянно выбраковывались наиболее подвижные или держащиеся далеко впереди или в стороне от основного стада. В результате отбора олени чукотской породы по поведению на пастбищах резко отличаются от других пород. У них сильнее развит инстинкт стадности, менее подвижны, приспособлены к пастбищам с невысоким содержанием кормовых лишайников. Биологические особенности оленей данной породы – это скороспелость, высокая степень нажировки, с ранним сроком гона и отела, приспособленные к тундровым зонам разведения.
Породе присуще низкорослость, с округленным туловищем. В отличие от других пород способны за кратчайший срок достигать высокую упитанность, что позволяет им лучше переносить суровые климатические условия и бескормицу во время гололеда. [33].
Доминирующая масть бурая, среди разводимых в Якутии – темно-бурая. Шерстный покров более богат, с заметной ярусностью, содержит много пуха. Остевые волосы более длиннее и гуще.
Более приспособлены к тундровой зоне, с прохладным коротким летним периодом, продолжительной малоснежной зимой. По сравнению с другими породами отел у них проходит на 15-20 дней раньше. Тугуты очень быстро растут и 4-5 месяцам способны нарастить большой запас питательных веществ.
Нижнеколымский район расположен на северо-востоке РС(Я), охватывает зону тундры и частично лесотундры. На севере граница Нижнеколымского района омывается водами ВосточноСибирского моря, на западе граничится с Аллаиховским районом, на юге со Среднеколымским районом, на востоке с Магаданской областью.
Площадь района составляет 9272974 га. За кооперативом «Турваургин» закреплено в долгосрочное пользование 1908000 га, земельных угодий, в том числе оленьих пастбищ – 1526786 га земли.
Климат Нижнеколымского района среднеконтинентальный. Здесь сказывается влияние западных циклонов, приносящих тепло и влагу, что вызывает значительное потепление. Благодаря этому температура зимних месяцев в районе значительно выше, чем в центральной Якутии. Среднегодовая температура – 11,6 градусов. Переход к положительным температурам совершается в последних числах мая, к отрицательным – в конце сентября. Заморозки прекращаются в конце июня и наступают во второй декаде августа. В отдельные годы заморозки наблюдаются уже в июле.
Годовое количество осадков составляет в среднем 285 мм. Более 70% осадков выпадает летом. Общее количество дней с осадками за год составляет 138-235.
Ветры на побережье имеют муссонный характер, то есть с сентября по март большей частью господствуют южные ветры, направленные в сторону полярного бассейна. В летнее время они дуют в обратном направлении с моря на сушу. Число дней с сильными ветрами подвержено большим колебаниям и составляет от 1 до 54. Сильные ветры часто наблюдаются на побережье моря. Малое число дней с сильными ветрами характерно для лесотундры.
Ветер имеет большое значение для оленеводства и содержания оленей. В летнее время, особенно в июле, наблюдается большое количество насекомых, из-за которых олени плохо используют корма и теряют упитанность.
На побережье в июле – начало августа скорость ветра составляет 4-7 м/сек. Ветер способствует благоприятному выпасу оленей, поэтому лучшие летние оленьи пастбища расположены вдоль побережья.
Зимой ветры дуют с материка, неся охлажденные массы воздуха. В январе скорость ветра в тундре составляет 5-7 м/сек., а в лесотундре – 3,2-3,6 м/сек. Сильные ветры имеют исключительное значение для распределения снежного покрова, а также влияют на растительность. Они местами уносят с поверхности земли не только снег, но и минеральные частицы, местами уплотняют снег, местами создают снежные заносы, укрывающие растения в зимний период.
Сильные ветры способствуют большому испарению влаги, что, при недостатке осадков и тепла, отрицательно сказывается на растениях. В целях защиты растения приобретают приземистую, часто подушкообразную форму.
На побережье Северного Ледовитого океана с высокой относительной влажностью туманы наблюдаются часто и очень устойчиво, особенно в теплый период года. При этом максимум туманов отмечается в июле и минимум в декабре-январе.
Большим количеством дней с метелями отмечается тундра, особенно в январе-феврале.
На территории Нижнеколымского района, где зима длится 8 месяцев, в течение года выпадает 30-50% осадков в твердом виде. Снеговой покров не редко появляется в августе. Устойчивый же снеговой покров образуется во второй или третьей декаде сентября. Высота его постепенно увеличивается, и максимальный – достигает в первой половине апреля.
Мощность снегового покрова составляет 30-50 см. Число дней со снеговым покровом составляет 242-249. Уменьшение снегового покрова начинается в тундре с третьей декады апреля. В первой декаде июня субарктическая тундра освобождается от снега.
Наибольшая плотность снега в районе наблюдается на открытой местности, подверженной сильным ветром. Плотность снега, как и его высота, увеличивается в течении зимы.
Оленьи пастбища охватывают всю территорию Нижнеколымского района, практически не используются только бескормные участки и частично горы.
Размещение и фактически сложившееся использование и сезонных пастбищ совпадает с природными растительными зонами и подзонами, так, летние пастбища занимают территорию арктических тундр, примыкающую на севере к побережью ВосточноСибирского моря. Южнее летовок расположены пастбища переходных сезонов (поздней весны и ранней осени), занимающие северную и среднюю части субарктических тундр. Здесь наряду с зелеными кормами имеются лишайники, которые необходимы в эти периоды.
Дальше на юг количество ягеля увеличивается, покрытие его на отдельных участках доходит до 15-30%. Поэтому южная часть субарктических тундр используется как ранневесенние и позднеосенние пастбища.
Зимние пастбища расположены, в основном, в лесной зоне. Здесь наличие ягеля и природные условия вполне благоприятствуют зимнему выпасу оленей. Обеспеченность кормами хорошая и имеется защита от непогоды.
Движение оленьих стад в течение года происходит с юга на север и обратно. В летнее время стада находятся ближе к морю, в безлесной тундре, что обеспечивает нормальные условия выпаса в жаркий и комарный период. С наступлением прохладных дней стада поворачиваю на юг и, постепенно двигаясь, приходят на зимовку в лесную зону. С половины зимы стада вновь поворачивают на 180 градусов и двигаются уже в северном направлении на отельные и затем на летние пастбища.
Специально выделенных проходных путей на переходных пастбищ нет, так как сезонные пастбища контактируют между собой.
В таблицах 10 и 11 показаны линейные промеры оленей чукотской породы разводимых в СПК КРО «Турваургин» Нижнеколымского района. У самцов высота в холке составило 110,2 см, у самок 103,3 см, высота в локте составило у самцов 63,2 см, у самок 59,3 см, косая длина туловища у самцов 109,5 см, самки 109,3 см, обхват груди у самцов – 126,1 см, у самок – 125,0 см, обхват пясти у самцов 13,8 см, у самок – 11,7 см. (таблица 11,12).
Таблица 11–Линейные промеры самцов (хоры) (n-64 гол)
| Показатели | средний | минимальный | максимальный |
| Высота в холке, см. | 110,2 | 103,0 | 116,0 |
| Косая длина туловища, см | 109,5 | 102,0 | 121,0 |
| Высота в локте, см | 63,2 | 60,0 | 70,0 |
| Обхват груди, см | 126,1 | 120,0 | 130,0 |
| Обхват пясти, см | 13,8 | 13,0 | 14,0 |
Таблица 12 – Линейные промеры самок (важенок) (n-42 гол)
| Показатели | средний | минимальный | максимальный |
| Высота в холке, см. | 103,3 | 90,0 | 112,0 |
| Косая длина туловища, см | 109,3 | 98,0 | 120,0 |
| Высота в локте, см | 59,3 | 55,0 | 65,0 |
| Обхват груди, см | 125,0 | 115,0 | 133,0 |
| Обхват пясти, см | 11,7 | 11,0 | 12,0 |
При определении особенностей телосложения оленей чукотской породы использовали индекс телосложения, соотношением отдельных промеров, выраженных в процентах. Данные представлены в таблице 12.
Таблица 13 – Индексы телосложения чукотской породы оленей
| Индексы, % | СПК КРО «Турваургин» | По литературным данным | ||
| самцы | самки | самцы | самки | |
| Растянутости | 99,3 | 105,8 | 106,5 | 108,7 |
| Костистости | 12,5 | 11,3 | 12,9 | 11,7 |
| Массивности | 114,4 | 121,1 | 121,3 | 119,7 |
| Длинноногости | 57,4 | 57,4 | 57,1 | 58,8 |
| Сбитости | 115,2 | 114,4 | 113,9 | 110,1 |
Исходя из данных индексов телосложения олени чукотской породы по сравнению с другими породами более массивны, индекс сбитости равняется у самцов 115,2 см, у самок 114,4 см. Костяк хорошо развит – индекс костистости самцов составил 12,5 см, самок 11,3 см, что характерно для оленей с хорошо выраженным мясным типом.
У оленей СПК КРО «Турваургин» средняя живая масса у самцов чукотской породы – 103,3 кг, у самок – 94,5 см.
Таблица 14 – Живая масса самцов чукотской породы оленей, кг
| Показатели | по литературным данным | (n-64 гол) | ||
| средний | min | max | ||
| Живая масса, кг | 107,0 | 103,3 | 98,0 | 112,0 |
Таблица 15 – Живая масса самок чукотской породы оленей, кг
| Показатели | по литературным данным | (n-42 гол) | ||
| средний | min | max | ||
| Живая масса, кг | 94,5 | 84,3 | 68,0 | 99,0 |
2.4. Генетическая структура пород домашних северных оленей
По результатам молекулярно-генетического тестирования установлено, что микросателлитные маркеры имеют широкий спектр аллелей. Общее количество выявленных аллелей составил 127, и варьировало от 2 (C217) до 14 (BMS1788). В среднем на локус приходилось 7,9 аллели (Na), среднее число эффективных аллелей (Ne) составил 4,1 (таблица16).
Показатель индекса фиксации отрицательный в 7 локусах из 16, от -0,006 в локусе OheQ (Na=12) до -0,103 в локусе T40 (Na=9). В локусах Rt6, BMS1788, Rt9, C143, Rt7, C217, Rt24, BMS745 и NVHRT16 индекс фиксации положительный, максимальное его значение выявлено в локусе C143 (F=0,125), среднее значение по породе 0,001.
Таблица 16 – Полиморфизм 16 микросателлитных маркеров у оленей чукотской породы (n=90)
| Локус | Na | Ne | Ho | He | F | PIC |
| Rt6 | 8 | 4,519 | 0,744 | 0,779 | 0,044 | 0,750 |
| BMS1788 | 14 | 6,252 | 0,789 | 0,840 | 0,061 | 0,822 |
| Rt 30 | 10 | 5,261 | 0,867 | 0,810 | -0,070 | 0,787 |
| Rt1 | 11 | 3,824 | 0,778 | 0,739 | -0,053 | 0,787 |
| Rt9 | 10 | 4,099 | 0,744 | 0,756 | 0,015 | 0,716 |
| C143 | 3 | 2,142 | 0,467 | 0,533 | 0,125 | 0,427 |
| Rt7 | 7 | 3,689 | 0,711 | 0,729 | 0,024 | 0,696 |
| OheQ | 12 | 6,678 | 0,856 | 0,850 | -0,006 | 0,833 |
| FCB193 | 8 | 2,980 | 0,667 | 0,664 | -0,003 | 0,628 |
| C217 | 2 | 1,328 | 0,222 | 0,247 | 0,101 | 0,217 |
| Rt24 | 10 | 5,678 | 0,811 | 0,824 | 0,016 | 0,803 |
| C32 | 3 | 2,973 | 0,711 | 0,664 | -0,072 | 0,590 |
| BMS745 | 6 | 4,519 | 0,778 | 0,779 | 0,001 | 0,743 |
| NVHRT16 | 8 | 5,542 | 0,811 | 0,820 | 0,010 | 0,795 |
| T40 | 9 | 2,527 | 0,667 | 0,604 | -0,103 | 0,573 |
| C276 | 6 | 3,596 | 0,778 | 0,722 | -0,077 | 0,946 |
| Среднее | 7,94 | 4,101 | 0,713 | 0,710 | 0,001 | 0,695 |
Наблюдаемая гетерозиготность (Ho) составила 0,713, ожидаемая (He) 0,710, Но>He — система случайного скрещивания преобладает над инбридингом. Минимальное значение наблюдаемой гетерозиготности выявлена в локусе C217, имеющем 2 аллели, и составила 0,222, при этом индекс фиксации 0,101.
Индекс полиморфизма (PIC) варьировал от 0,217 до 0,946, в среднем 0,695. Согласно формулировке Botstein et. al. (1980) Слегка информативным (PIC<0.25) оказался один локус — C217, также локус C143 — достаточно информативным (0.5>PIC >0,25). Четырнадцать локусов шестнадцати показали высокую информативность (PIC>0,5), при этом у 10 локусов индекс информативности выше 0,7.
В результате исследования полиморфизма 16 локусов микросателлитной ДНК у оленей чукотской породы завезенных с Чукотского АО (МП СХП БМР «Островное») обнаружено, что они имеют определённый спектр аллелей и своеобразный генетический профиль. Общее количество выявленных аллелей составило 116. Число эффективных аллелей составил 3,92. В среднем на 1 локус приходиться 7,25 аллеля. Индекс фиксации показывает на избыток гетерозигот и составил – 0,030 (таблица 17).
Таблица 17 — Полиморфизм 16 микросателлитных маркеров у оленей чукотской породы завезенных с Чукотского АО (n=54)
| Локус | Na | Ne | Ho | He | F |
| Rt6 | 8 | 4,643 | 0,852 | 0,785 | -0,086 |
| BMS1788 | 13 | 4,707 | 0,778 | 0,788 | 0,012 |
| Rt 30 | 9 | 4,636 | 0,852 | 0,784 | -0,086 |
| Rt1 | 9 | 3,317 | 0,704 | 0,699 | -0,007 |
| Rt9 | 9 | 4,840 | 0,870 | 0,793 | -0,097 |
| C 143 | 2 | 1,890 | 0,500 | 0,471 | -0,062 |
| Rt7 | 7 | 3,962 | 0,667 | 0,748 | 0,108 |
| OheQ | 12 | 7,439 | 0,833 | 0,866 | 0,037 |
| FCB193 | 8 | 3,331 | 0,796 | 0,700 | -0,138 |
| C217 | 3 | 1,275 | 0,204 | 0,216 | 0,055 |
| Rt24 | 8 | 4,662 | 0,778 | 0,785 | 0,010 |
| C32 | 3 | 2,760 | 0,593 | 0,638 | 0,071 |
| BMS745 | 6 | 4,382 | 0,852 | 0,772 | -0,104 |
| NVHRT16 | 8 | 5,166 | 0,796 | 0,806 | 0,013 |
| T40 | 6 | 1,889 | 0,519 | 0,471 | -0,102 |
| C276 | 5 | 3,777 | 0,815 | 0,735 | -0,108 |
| Среднее | 7,25 | 3,917 | 0,713 | 0,691 | -0,030 |
Сравнительная оценка хоров производителей двух популяций показал, умеренное генетическое расстояние между изученными животными и составил 0,035 ед. (таблица 18).
Таблица 18 – Сравнительная оценка хоров производителей
| n | Na | Ne | F | PA | NeiD | |
| МП СХП БМР «Островное» | 54 | 7,250 | 3,917 | -0,030 | 11 | 0.035 |
| СПК КРО «Турваургин» | 55 | 7,563 | 4,114 | 0,019 | 16 |
Показатель индекса фиксации у хоров производителей СПК КРО «Турваургин» положительный, что говорит о дефиците гетерозиготных особей. По среднему количеству аллелей на локус числу эффективных аллелей хоры СПК КРО «Турваургин» превосходят хоров производителей МП СХП БМР «Островное».
При сравнительной оценке оленей чукотской породы (местных и привезенных с Чукотского АО), у хоров СПК КРО «Турваургин» идентифицировано 11 приватных аллелей. В локусе BMS1788 обнаружено три аллеля с числом повторностей 10, 12,1 и 15,1 частота встречаемости которых составила 0,009; 0,028 и 0,019 соответственно. Аллель локуса FCB193 с числом повторностей 12 встречалась с частотой 4,6% (таблица 19).
Таблица 19 – Частота встречаемости приватных аллелей у оленей чукотской породы, разводимых на территории республики Саха (Якутия).
| Локус | Аллель | Частота |
| BMS1788 | 10 | 0,009 |
| BMS1788 | 12,1 | 0,028 |
| BMS1788 | 15,1 | 0,019 |
| Rt1 | 22 | 0,019 |
| Rt9 | 19 | 0,009 |
| FCB193 | 12 | 0,046 |
| C217 | 8,1 | 0,019 |
| Rt24 | 16 | 0,009 |
| BMS745 | 15 | 0,009 |
| NVHRT16 | 20 | 0,009 |
| T40 | 27,3 | 0,019 |
В 2019 году в стаде №9 хоры производители были полностью заменены закупленными хорами с МП СХП БМР «Островное». Оценка генетического разнообразия оленей чукотской породы после замены хоров производителей представлена в таблице 20.
Таблица 20 – Оценка генетического разнообразия оленей чукотской породы
| n | Na | Ne | Ho | He | F | |
| СПК КРО «Турваургин» | 89 | 8,00 | 4,016 | 0,718 | 0,700 | -0,027 |
В результате замены хоров индекс фиксации в популяции показывает избыток гетерозиготных особей (F=-0,027). Наблюдаемая гетерозиготность составила 0,718 и ожидаемая гетерозиготность 0,700.
У оленей эвенской породы всего выявлено 122 аллеля. В среднем на локус приходилось 7,625 аллеля. Наибольшее число аллелей наблюдалось в локусе BMS2088 (Na=14), а минимальное количество (Na=2) в локусах C143 и C2188. Число эффективных аллелей варьировало 1,325 (C2188) до 7,463 (OheQ). Индекс фиксации принимал положительное значение в 7 локусах: BMS2088, Rt1, Rt9, Rt7, OheQ, RT24 и NVHRT16. В среднем индекс фиксации составил -0,021, что указывает на избыток гетерозиготных особей.
Таблица 21 – Полиморфизм 16 микросателлитных маркеров у оленей эвенской породы (n=178)
| Локус | Na | Ne | Ho | He | F |
| RT6 1 | 10 | 3,228 | 0,697 | 0,690 | -0,009 |
| BMS2088 | 14 | 3,519 | 0,685 | 0,716 | 0,042 |
| RT30 | 7 | 4,085 | 0,758 | 0,755 | -0,004 |
| RT1 | 10 | 5,112 | 0,775 | 0,804 | 0,036 |
| RT9 | 9 | 5,369 | 0,792 | 0,814 | 0,027 |
| C143 | 2 | 1,926 | 0,579 | 0,481 | -0,204 |
| RT7 | 9 | 5,461 | 0,792 | 0,817 | 0,030 |
| OHEQ | 12 | 7,463 | 0,860 | 0,866 | 0,007 |
| FCB1533 | 9 | 4,474 | 0,837 | 0,776 | -0,078 |
| C2188 | 2 | 1,325 | 0,253 | 0,245 | -0,030 |
| RT24 | 10 | 6,156 | 0,792 | 0,838 | 0,054 |
| C32 | 3 | 2,531 | 0,635 | 0,605 | -0,049 |
| BMS745 | 6 | 2,919 | 0,669 | 0,657 | -0,017 |
| NVHRT16 | 7 | 3,841 | 0,736 | 0,740 | 0,005 |
| T40 | 7 | 1,643 | 0,410 | 0,391 | -0,048 |
| C276 | 5 | 3,117 | 0,742 | 0,679 | -0,092 |
| Среднее | 7,625 | 3,886 | 0,688 | 0,680 | -0,021 |
Результаты расчетов показали, что в 8 локусах из 16 наблюдается недостаток гетерозигот, максимальное его значение составлял 0,042 в локусе BMS2088. В среднем по породе индекс фиксации (F) составил -0,021, что указывает избыток гетерозиготных особей. Наблюдаемая гетерозиготность (Ho) составила 0,688, ожидаемая (He) 0,680, Но>He — система случайного скрещивания преобладает над инбридингом (таблица 21).
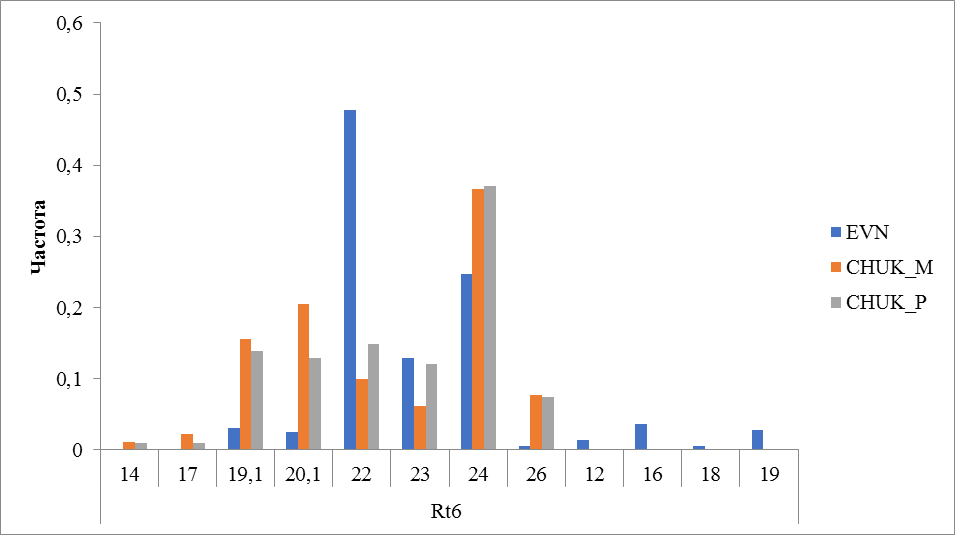
Рисунок 3 Частота встречаемости аллелей локуса Rt6
В локусе Rt6 (рисунок 3) выявлено 12 аллелей, у оленей эвенской породы обнаружена – 10, наиболее часто встречался аллель с числом повторности 22 (0,478). У оленей чукотской породы выявили 8 аллелей, как у местных так у привозных оленей преобладал аллель с числом повторности 24 (37%).
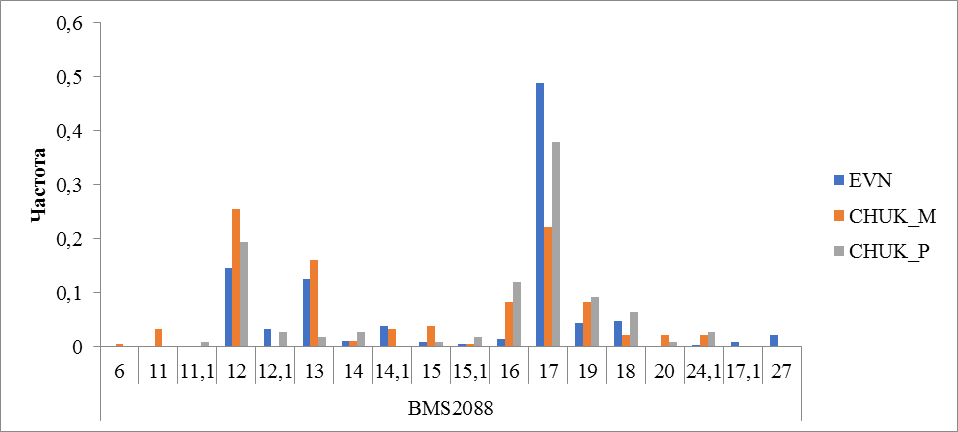
Рисунок 4 Частота встречаемости аллелей локуса BMS2088
Аллельное разнообразие локуса BMS2088 представлена 18 аллелью. У оленей эвенской породы выявлено 14 аллелей, 13 и 15 у оленей чукотской породы привозных и местных соответственно. Частота встречаемости аллеля с числом повторности 17 составила 0,489 в эвенской породе, 0,380 и 0,256 у привозных и местных оленей чукотской породы, соответственно (рисунок 4).
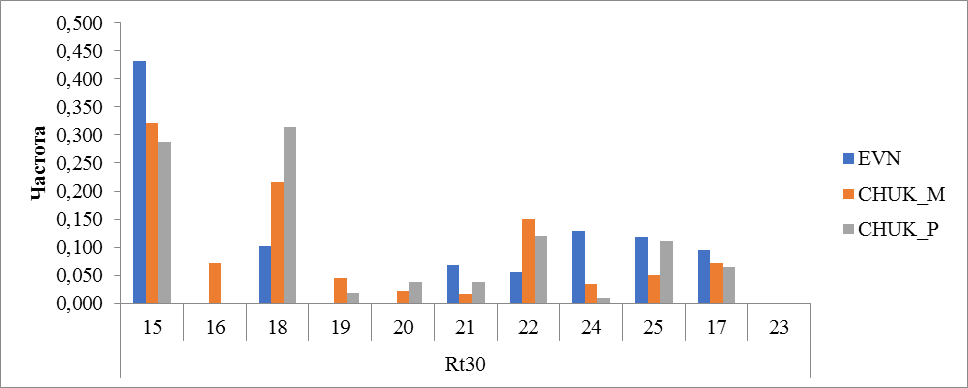
Рисунок 5 Частота встречаемости аллелей локуса Rt30
В генотипе исследованных пород оленей локус Rt30 имел 11 аллелей. У оленей эвенской породы выявлено 8. Аллель с количеством повторов 15 встречался с частотой 0,433 у оленей эвенской породы. У оленей чукотской породы частота этого аллеля составила 0,322 у местных и 0,287 у привозных. У оленей чукотской породы привезенных с Чукотского АО преобладали аллели с числом повторности 18 (0,315) и 15(0,287) (рисунок 5).
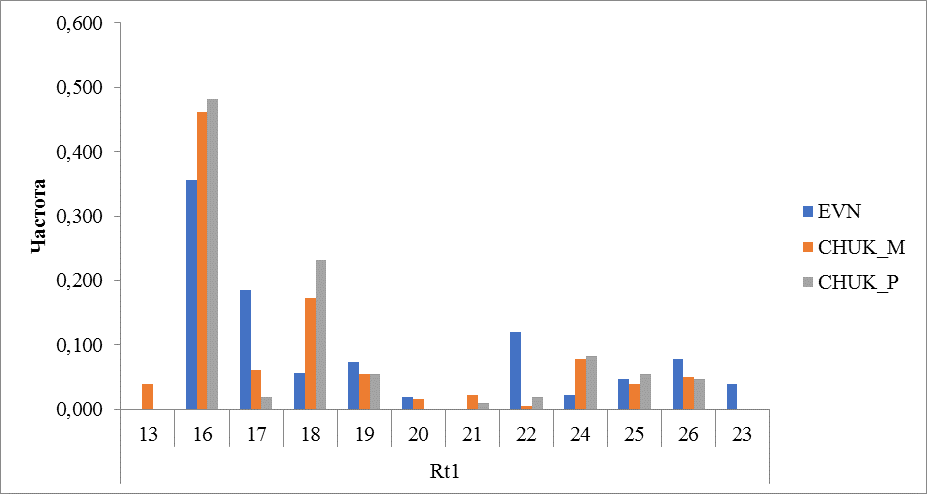
Рисунок 6. Частота встречаемости аллелей локуса Rt1
Микросателлитный локус Rt1 представлен 12 аллелями. Во всех исследованных группах (породах) преобладал аллель с количеством повторов 16. Частота встречаемости его составила 0,357 в эвенской породе, 0,461 и 0,481 у местных и привозных оленей чукотской породы соответственно. Наибольшее количество аллелей выявлено у местных оленей чукотской породы – 11, отсутствует аллель с числом повторности 23, которая присутствует в эвенской породе. Этот аллель также не выявлен у привозных оленей чукотской породы. Между привозными и местными оленями этой породы также имеется отличие в аллельном разнообразии. Так у привозных оленей генотипировано 9 аллелей. Не обнаружены аллели с количеством повторов 13 и 20 (рисунок 6).
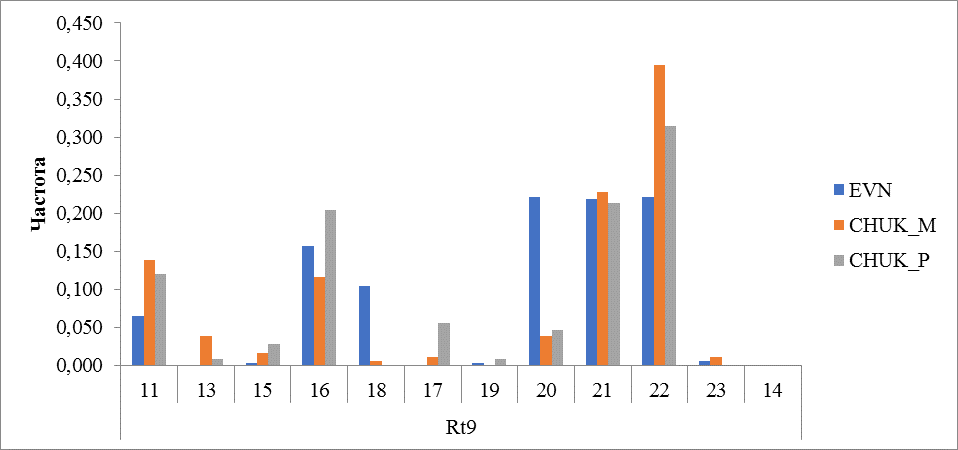
Рисунок 7 Частота встречаемости аллелей локуса Rt9
У протестированных оленей в локусе Rt9 было обнаружено 11 аллелей. Наибольшую частоту встречаемости от 0,222 до 0,394 во всех исследованных группах (порода) имел аллель 22, на втором месте по частоте встречаемости занимал аллель 21 (0,213-0,228).
Аллель с количеством повторов 19 встречался меньше всего у оленей эвенской породы (0,003) и у привозных оленей чукотской породы (0,009). У местных чукотских оленей этот аллель не обнаружен. В этом локусе также выявлено породное отличие по аллельному разнообразию. Аллели 13 и 17, у которых частота встречаемости у оленей чукотской породы составила от 0,009 до 0,039, не обнаружена у оленей эвенской породы. У оленей эвенской породы меньше всего встречались аллели 15 и 19 с частотой 0,003 (рисунок 7).
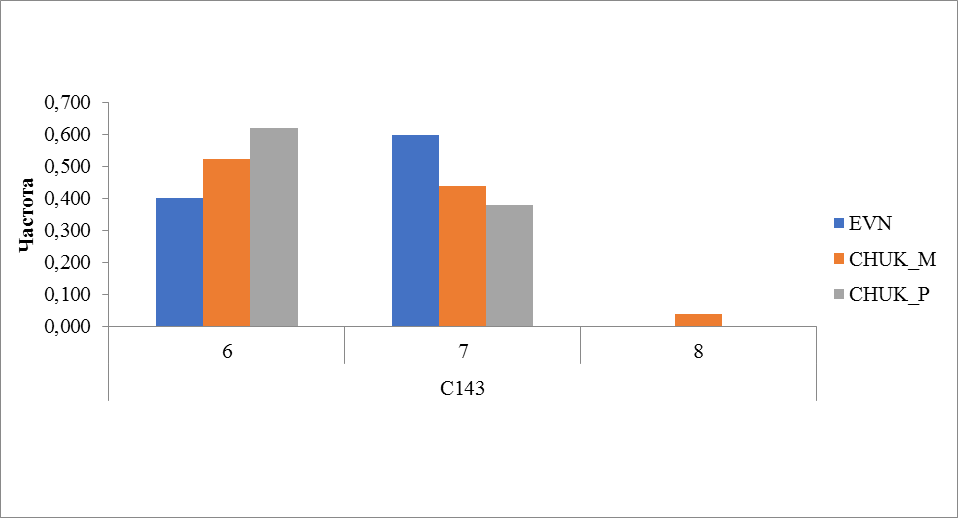
Рисунок 8 Частота встречаемости аллелей локуса C143
Микросателлитный локус C143 (рисунок 8) у домашних северных оленей представлен 3 аллелями. Аллель с числом повторности 8 выявлена у местных оленей чукотской породы, его частота встречаемости составила 0,039. В этом локусе отмечено межпородное различие, так если у оленей чукотской породы преобладал аллель с количеством повторов 7, то у оленей эвенской породы наиболее часто встречался аллель 6 (0,598).
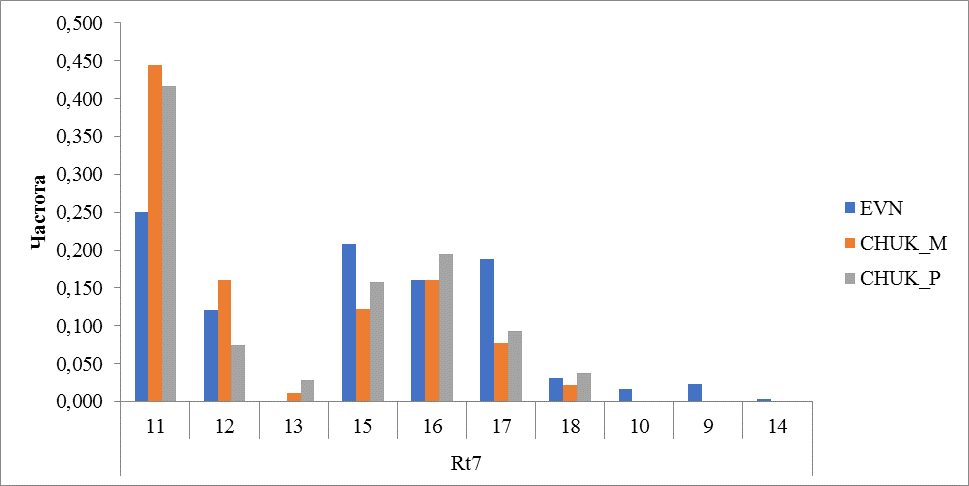
Рисунок 9 Частота встречаемости аллелей локуса Rt7
Заметное межпородное отличие выявлено по локусу Rt7. У оленей эвенской породы выявлено 9 аллелей, чаще встречались аллели 11 (0,250) и 15 (0,208). У оленей чукотской породы обнаружено 7 аллелей, как у местных, так и у привозных оленей наиболее часто встречался аллель с количеством повторов 11 (0,444 и 0,417 соответсвенно). У оленей чукотской породы отсутствовали аллели 9, 10 и 14, но зафиксирован аллель 13, который не обнаружен у оленей эвенской породы (рисунок 9).
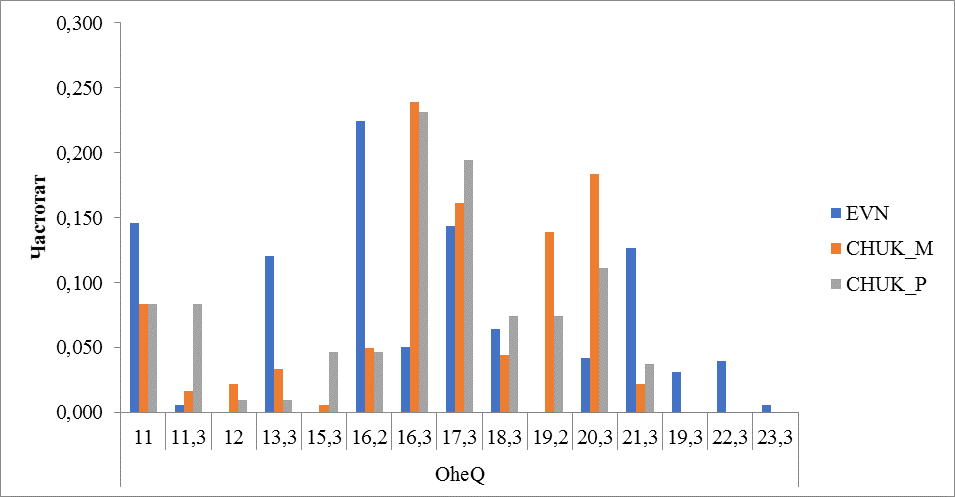
Рисунок 10 Частота встречаемости аллелей локуса OheQ
Высокое генетическое разнообразие (15 локусов) было выявлено в локусе OheQ. При этом в каждой группе (породе) выявлено по 12 аллелей, но с заметным отличием по аллельному разнообразию. У протестированных оленей эвенской породы отсутствовали алелли с числом повторности 12, 15,3 и 19,2. Наиболее часто встречался аллель 16,2 (0,225) когда как у оленей чукотской породы этот аллель встречался с частотой 0,050 и 0,046 у местных и привозных соответственно. Олени чукотской породы охарактеризовались преобладанием дробного аллеля 20,3 (0,183 у местных и 0,111 у привозных), а у оленей эвенской породы этот аллель встречался с частотой 0,042 (рисунок 10).
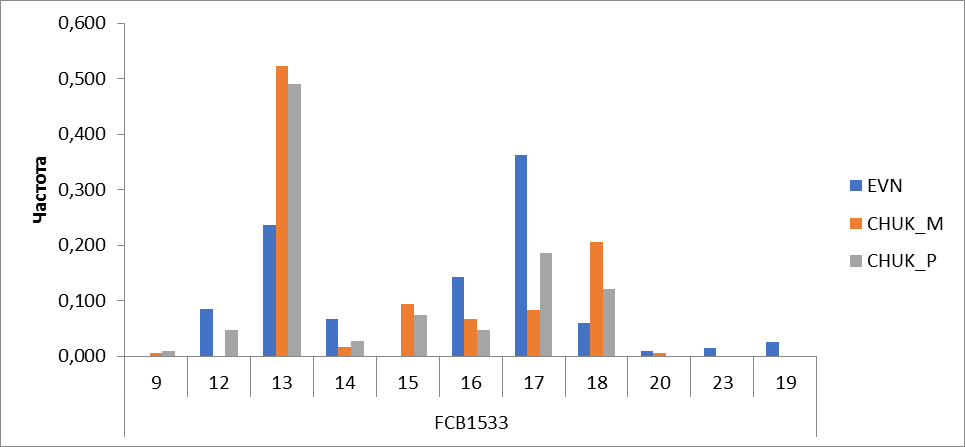
Рисунок 11 Частота встречаемости аллелей локуса FCB1533
В локусе FCB1533 протипировано 11 аллелей. В эвенской породе выявлено 9 аллелей, наиболее распространенными аллелями в породе являются аллели 17 (0,362) и 13 (0,236). Аллель с количеством повторов 13 у оленей чукотской породы преобладал больше всего, и частота встречаемости составила у местных – 0,522 и 0,491 у привозных. Эвенская порода отличалась отсутствием аллелей 9 и 15. Между местными и привозными оленями также имелось отличие. У оленей, привезенных с Чукотского АО, присутствует аллель 12 (0,046), а у местных этот аллель не был обнаружен, и наоборот аллель с числом повторности 20 выявленный у местных отсутствовал у привозных (рисунок 11).
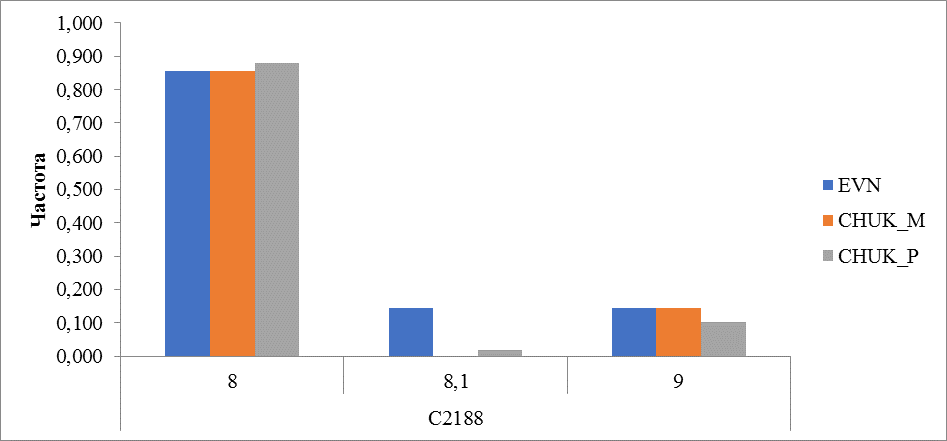
Рисунок 12 Частота встречаемости аллелей локуса C2188
В исследованных породах микросателлитный локус C2188 имел 3 аллеля и характеризуется преобладанием аллеля с количество повторов 8. В эвенской породе частота встречаемости этого аллеля достигала 0,857, у местных чукотских оленей 0,856 и 0,880 у привозных. У местных оленей чукотской породы отсутствовал дробный аллель 8,1, который встречался у оленей эвенской породы с частотой 0,143 и 0,019 у привозных оленей чукотской породы. Частота встречаемости аллеля с числом повторности 9 варьировала от 0,102 до 0,145 (рисунок 12).
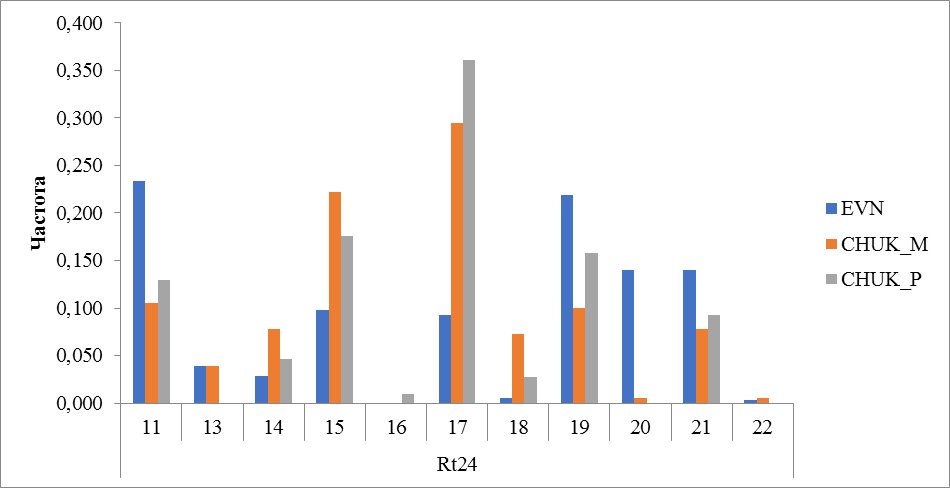
Рисунок 13 Частота встречаемости аллелей локуса Rt24
В эвенской породе и у местных оленей чукотской породы выявлено 10 аллелей из 11 идентифицированных в локусе Rt24. Аллель с количеством повторов 16 отсутствовал в этих группах (породах), частота встречаемости этого аллеля у закупленных оленей чукотской породы составила 0,009. Помимо этого, привозные олени чукотской породы отличились отсутствием в генотипе аллелей 13 и 22. В целом по чукотской породе наиболее распространенным являлся аллель 17, частота встречаемости у местных составила 0,294 и 0,361 и привозных, а у оленей эвенской породы частота встречаемости этого аллеля составила 0,093. В эвенской породе преобладающее количество имеют аллели с числом повторности 11 (0,233) и 19 (0,219). К редким аллелям локуса Rt24 можно отнести аллель с количеством повторов 22, в эвенской породе частота встречаемости составила 0,003 и 0,006 у местных оленей чукотской породы (рисунок 13).
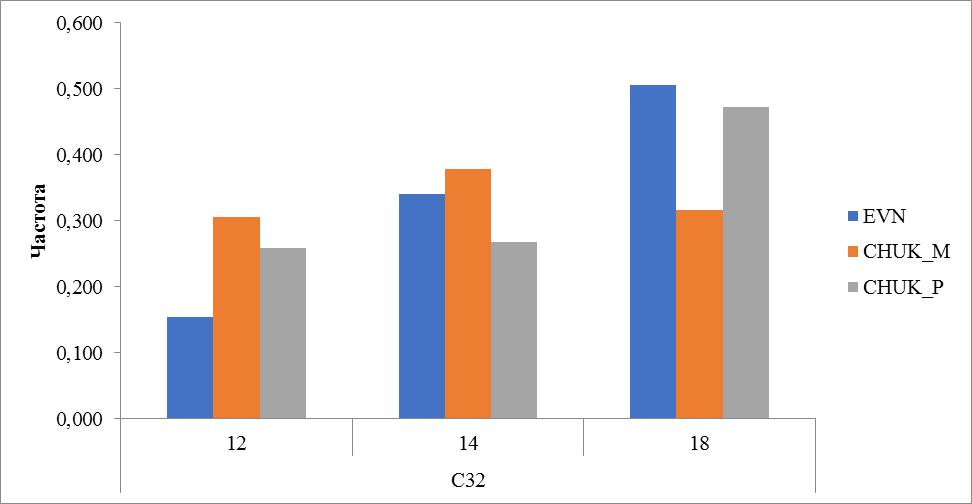
Рисунок 14 Частота встречаемости аллелей локуса С32
По локусу С32 определены три аллеля с количеством повторов 12, 14 и 18. Если у местных оленей чукотской породы частота встречаемости этих аллелей распределена более равномерно (0,306, 0,378 и 0,317) , то у привозных и оленей эвенской породы преобладающее большинство имеет аллель 18, частота встречаемости которого составила 0,472 и 0,506 соответственно (рисунок 14).
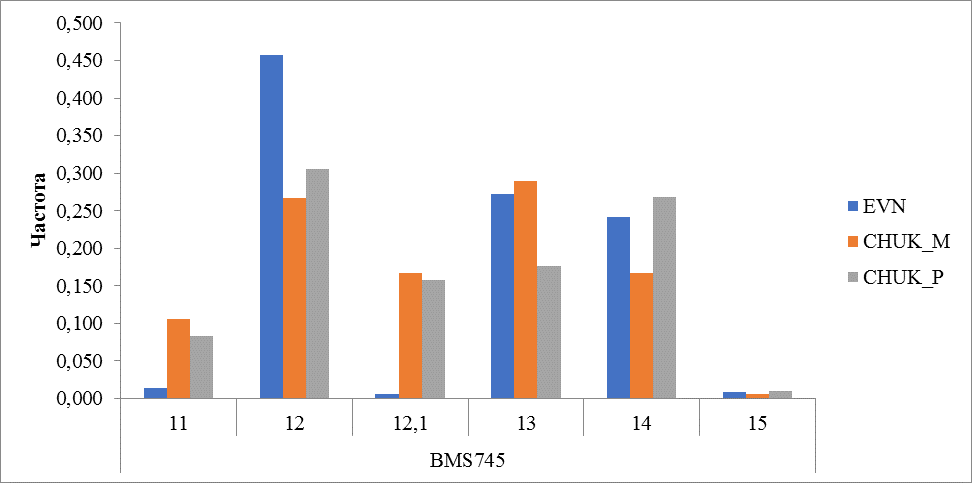
Рисунок 15 Частота встречаемости аллелей локуса BMS745.
В микросателлитном локусе BMS745 выявлено 6 аллелей. В эвенской породе на первом месте по частоте встречаемости расположился аллель 12 (0,458), редкими для этой породы оказались аллели 12,1 и 15 (0,006-0,005). У оленей чукотской породы наиболее часто встречались аллели 12, 13 и 14, аллель с количеством повторов 15 оказался редким (0,006-0,009) для чукотских оленей, как и у эвенских оленей (рисунок 15).
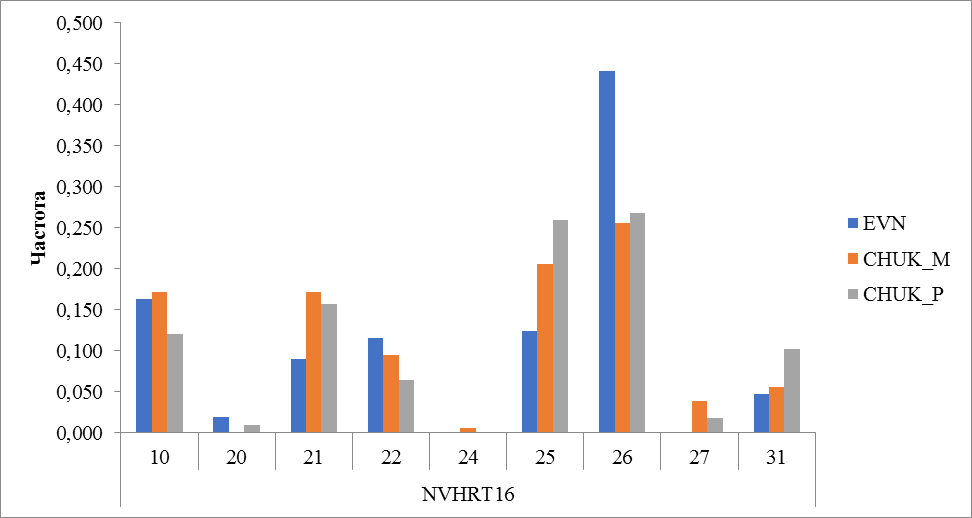
Рисунок 16 Частота встречаемости аллелей локуса NVHRT16.
По локусу NVHRT16 у оленей чукотской породы выявлено больше аллелей (8 из 9) по сравнению с эвенской породой (7). У оленей эвенской породы не обнаружены аллели 24 и 27. Аллель с количеством повторов 24 также не выявлен у привозных оленей чукотской породы. Частота встречаемости этого аллеля у местных оленей чукотской породы также невысока и составила 0,006. У местных чукотских оленей не обнаружен аллель 20, частота встречаемости его у оленей эвенской породы составила 0,020 и 0,009 у привозных оленей чукотской породы. Кроме этого, необходимо отметить, что аллель 26 преобладал по частоте встречаемости во всех исследованных группах (породах) и составила 0,441 в эвенской породе, 0,256 у местных и 0,269 у привозных оленей чукотской породы (рисунок 16).
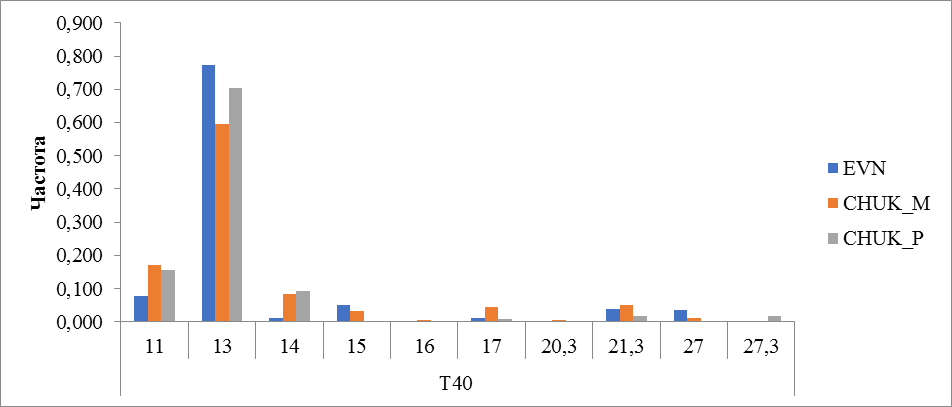
Рисунок 17 Частота встречаемости аллелей локуса T40
У исследованных животных выявлено 10 аллелей локуса T40. Наибольшее его количество обнаружено у местных оленей чукотской породы (9 аллелей), у оленей эвенской породы выявлено 7 аллелей и 6 у привозных оленей чукотской породы. При немалом количестве аллелей большая доля встречаемости принадлежит одному аллелю с числом повторности 13. Частота встречаемости этого аллеля у местных оленей чукотской породы равнялась 0,594, у оленей эвенской породы 0,772 и 0,704 у привозных оленей чукотской породы. Также олени чукотской породы отличались от эвенской по частоте встречаемости аллеля 11. В эвенской породе частота встречаемости аллеля 11 составила 0,079, когда как в чукотской породе от 0,157 до 0,172 (рисунок 17).
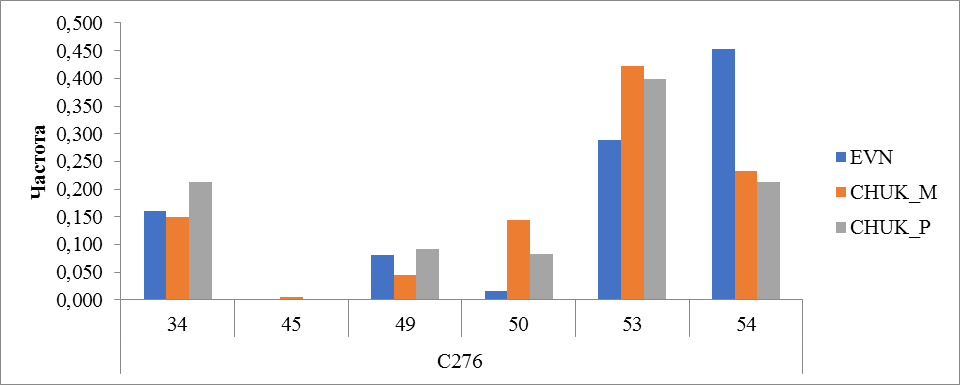
Рисунок 18 Частота встречаемости аллелей локуса C276
В локусе C276 выявлено 6 аллелей. У местных оленей чукотской породы выявлены все 6 аллелей, наиболее распространенным аллелем оказался аллель 53 (0,422). По частоте встречаемости этот аллель у привозных чукотских оленей также стоит на первом месте (0,398). У оленей эвенской породы чаще всего встречался аллель 54 (0,452). В этой породе и у привозных оленей чукотской породы не был обнаружен аллель 45. Частота встречаемости этого аллеля у местных оленей чукотской породы – 0,006 (рисунок 18).
Таблица 22 – Популяционно-генетическая характеристика популяций домашних северных оленей, разводимых на территории Республики Саха (Якутия)
| Показатели | Порода | ||
| Эвенская | Местная чукотская | Привозная чукотская | |
| Na | 7,625 | 7,938 | 7,250 |
| Na Freq. >= 5% | 4,438 | 4,750 | 4,563 |
| Ne | 3,886 | 4,101 | 3,917 |
| He | 0,680 | 0,710 | 0,691 |
*Примечание: Na — среднее число аллелей; Na Freq. >= 5% — среднее число аллелей имеющих частоту встречаемости больше или равно 5%; Ne — число эффективных аллелей; He-ожидаемая гетерозиготность
В таблице 22 представлены популяционно-генетические характеристики пород домашних северных оленей. На первом месте (Na=7.93, Ne=4.1) по числу аллелей на локус и по числу эффективных аллелей стоит чукотская порода оленей (местная). Также местные олени чукотской породы охарактеризовались наибольшим числом аллелей, имеющих частоту встречаемости больше или равно 5% и высоким уровнем ожидаемой гетерозиготности. Наименьшее количество эффективных аллелей наблюдалось у оленей эвенской породы, число аллелей, имеющих частоту встречаемости больше или равной 5%, составило 4,438. У привозных оленей чукотской породы число эффективных аллелей составило 3,917 при среднем числе аллелей на локус 7,250.
Таблица 23 – Генетическое расстояние между исследованными группами (породами) домашних северных оленей
| EVN | 0,000 | ||
| CHUK_М | 0,157 | 0,000 | |
| CHUK_Р | 0,130 | 0,032 | 0,000 |
| EVN | CHUK_М | CHUK_Р |
Анализ генетических расстояний по микросателлитам ДНК показывает минимальные различия (0,032) между местными оленями чукотской породы и оленями той же породы завезенных с Чукотского АО. Несмотря на разное происхождение и разную зону разведения наблюдалась относительно небольшая генетическая дистанция между оленями эвенской и чукотской пород. Наибольшее расстояние (0,130) наблюдалась с местными оленями чукотской породы (таблица 23).
Заключение
В результате проведённых исследований установлено, что каждая порода домашних северных оленей характеризуется определённым спектром частот аллелей по микросателлитным локусам и имеет свои особенности
Максимальное число выявленных аллелей который служит одним из показателей генетического разнообразия популяции был выявлен в чукотской породе и этот показатель составил 127 аллелей. Для чукотской породы оленей разводимой в СПК КРО «Турваургин» характерен высокий уровень гетерозиготности и в популяции система случайного скрещивания преобладает над инбридингом, несмотря на длительное разведение породы в Якутии без «обновления крови». За последние годы численность оленей чукотской породы резко снизилось. И такое снижение поголовья может ввести к потере разнообразия. Сохранение популяции оленей чукотской породы в Якутии является одним из важных задач республики, так как эта порода имеет хорошие мясные качества, обладает высокой жизнеспособностью и адаптирована в неблагоприятных тундровых условиях Якутии.
Разработанная, на основе этих исследований, база данных биологических, продуктивных особенностей и генетической структуры домашних северных оленей позволяет получить данные о внутри и межпородной дифференциации популяций и оценить степень генетического разнообразия породы, что в будущем даст возможность улучшить качество селекционно-племенной работы.
Выводы
- Установлено что микросателлитные маркеры имеют широкий спектр аллелей и в целом обладают высокой информативной ценностью для выявления генетических различий между животными и группами животных.
- Сравнительный анализ оленей чукотской породы показал умеренное генетическое расстояние между хорами, закупленными с Чукотского АО и местными СПК КРО «Турваургин, и составил 0,035 ед.
- Установлено смещение генетического равновесия в сторону дефицита гетерозиготных особей СПК КРО «Турваургин», и составил 0,001, что означает система случайного скрещивания, преобладает над инбридингом.
- Оценка генетического разнообразия оленей чукотской породы после замены хоров производителей, закупленными хорами, показывает на избыток гетерозигот, индекс фиксации составил F= -0,027.
- В результате генетического анализа оленей эвенской породы выявлено высокое генетическое разнообразие по микросателлитам ДНК. Среднее число аллелей на локус составил 7,6 и число эффективных аллелей – 3,9. Показатель наблюдаемой гетерозиготности преобладает над ожидаемой (Но>He), что означает система случайного скрещивания преобладает над инбридингом.
- В среднем индекс фиксации составил -0,021, что указывает на избыток гетерозиготных особей.
Список использованных источников
- Степанов А.И., Иванова Л.С., Павлова С.А., Пестерева Е.С., Захарова Г.Е., Даянова Г.И., Егорова И.К., Колесова М.С., Крылова А.Н., Антонова У.А., Халтанова Ф.Д., Попова Т.Р., Малышева В.Л., Ковлякова П.Е., Гурьева А.М., Багимова М.Ф., Попов И.И., Тимофеева М.С., Винокурова В.С., Николаева Ф.В. и др. Система ведения сельского хозяйства в Республике Саха (Якутия) на период 2016-2020 годы: методическое пособие. — К.: ООО Технопринт, 2017. — 416 с.
- Сыроватский Д.И. Организация и экономика оленеводческого производства. — Я.: — 2000. — 407 с.
- Сыроватский Д.И., Неустроев М.П. современное состояние и перспективы развития северного оленеводства в России: рекомендации. — М.: Росинформагротех. — 2007. — 132 с.
- Мухачев, А.Д. Морфологические особенности и мясная продуктивность северных оленей Полярного Урала: автореф. дис… канд. с.-х. наук / А.Д. Мухачев. — М.: -1968. — 20 с.
- Макушев, Ю.Е. Морфологические особенности и мясная продуктивность северных оленей Эвенкии (на примере оленей совхоза «Суриндинский»): автореф. дис. … канд. с.-х. наук / Ю.Е. Макушев. – М.:- 1976. — 28 с.
- Помишин, С.Б. Основы биометрии в оленеводстве. – Я.: — 1979. — 78 с.
- Мухачев, А.Д. Оленеводство: учебник. – М.: — 1992. — 122 с.
- Андреев, В.Н., Голосов И.М., Преображенский Б.В. Северные олени: — К.: -1972. — 172 с.
- Качан, А.П., Мухачев А.Д. Сравнительная морфологическая и продуктивная характеристика чукотских и эвенских оленей: научные труды / Сиб. отд-ние ВАСХНИЛ. – Н.: — 1980. — С. 38-45.
- Sharifzadeh A., Doosti A. Investigation of leptin gene polymorphism in Iranian native cattle // Bulgar. J. Vet. Med. — 2012. — V. 15. — No.2. — P. 86−92.
- Долматова И.Ю., Горелов П.В., Ильясов А.Д. Характеристика аллелофонда башкирской популяции симменталького скота по микросателлитам. Актуальные проблемы гуманитарных и естественных наук. — 2012.: — №2. — с. 52-54.
- Перчун А.В., Лазебная И.В., Перчун А.В., Белокуров С. Г. Монолокусные и полилокусные ДНК-маркеры в селекции крупного рогатого скота. сборник тезисов VI съезда Вавиловского общества генетиков и селекционеров (ВОГиС) и ассоциированные генетические симпозиумы. — Ростов-на-Дону. — 2014. — с. 175
- Крутикова А.Н., Дементьева Н.В., Митрофанова О.В., Позовникова М.В., Гончаров В.В. Полиморфизм гена гормона роста северных оленей. Генетика и разведение животных, 2016. — №2. — с. 8-12
- Харзинова В.Р., Доцев А.В., Соловьева А.Д., Федоров В.И., Охлопков И.М., Виммерс К., Рейер Х., Брем Г., Зиновьева Н.А. Популяционно-генетическая характеристика домашнего северного оленя в Республике Якутия на основании полногеномного SNP анализа. Сельскохозяйственная Биология, 2017. — том 52. — № 4. — с. 669-678 doi: 10.15389/agrobiology.2017.4.669rus
- Митрофанова О.В., Дементьева Н.В., Зозуля О.К., Гончаров В.В. Оценка возможности использолвания микросателлитных маркеров у северного оленя rangifer tarandus. Проблемы освоения и сохранения Арктики, материалы Всерос. Научн. Практ. конф. С117-118
- Cervini M. Genetic variability of 10 microsatellite markers in the characterization of Brazilian Nellore cattle (Bos indicus) / M. Cervini, F.Henrique-Silva, N. Mortari, E.Matheucci// Jr. Genet Mol Biol.- 2006.-29(3).-P.486-490.
- Carolino I. Implementation of a parentage control system in Portuguese beef-cattle with a panel of microsatellite markers /I. Carolino, O. Conceição e.a.// Genet Mol Biol.- 2009.-Vol.32 (2).-P. 306-311.
- Stevanovic, J. Maletic Evaluation of 11 microsatellite loci for their use in paternity testing in Yugoslav Pied cattle (YU Simmental cattle) /J.Stevanovic, Z. Stanimirovic, V. Dimitrijevic, M. / Czech J. Anim. Sci..- 2010.-Vol.55(6).-P. 221–226.
- Karima F. Mahrous. Genetic variations in horse using microsatellite markers/ Mahrous F. Karima, M. Hassanane, M. Abdel Mordy, Heba I. Shafey, Nagwa Hassan// Journal of Genetic Engineering and Biotechnology. – 2011. – B.9.- P.103-109.
- Breen M. Genetical and physical assignments of equine microsatellits- first integration of anchored markers in horse genome mapping / M. Breen, G. Lindgren, G. Binns et al. // Mammalian Genome. 1997. — Vol.8. — P. 267-273.
- Cervini, M. Genetic variability of 10 microsatellite markers in the characterization of Brazilian Nellore cattle (Bos indicus)/ M.Cervini, F.Henrique-Silva, N. Mortari, E. Matheucci Jr // Genet. Mol. Biol. – 2006-vol.29, N3 (http://www.scielo.br)
- Pevzner P.A. An Eulerian path approach to DNA fragment assembly/ Pevzner P.A., Tang H., Waterman M.S.// Proc. Natl. Acad. Sci. USA.-2001.- 98(17).- P.9748-9753.
- Compeau, Ph. E. C. Genome reconstruction: a puzzle with a billion pieces / Ph. E. C. Compeau, P.A. Pevzner // Bioinformatics for Biologists: Cambridge Books Online (ebook.cambridge.org) — 2009.: -p.36-65.
- Долматова И.Ю. Генетическая дифференциация внутрипородных линий уток по данным рестрикционного анализа митохондриальной ДНК / И.Ю. Долматова, И.Н. Ганиева // Вестник Башкирского университета. — 2006.: — №3. — с. 62-65.
- Фисинин В.И. Использование биохимических и молекулярно-генетических маркеров для оценки генетического разнообразия промышленных популяций уток. Рекомендации //В.И. Фисинин, И.Ю. Длматова, Р.Р. Гадиев. — У.: — БГАУ. — 2006. — 32 с.
- Столповский Ю.А., Лапшин А.В., Кол Н.В., Сулимова Г.Е., Глазко В.И. Полиморфизм молекулярно-генетических маркеров у овец романовской породы // Известия ТСХА выпуск 2, 2008. – с. 131-136.
- Ельсукова И.А., Феофилоф Ю.А., Юлдашбаев Ю.А., Глазко В.И Генетическая дифференциация суюндукского и бирликского внутрипородных типов эдильбаевской породы овец. // Известия ТСХА выпуск 6, 2010. – с. 84-89
- Проскурина Н.В. Сравнительный анализ информативности эритроциатрных антигенов и ДНК микросателлитов как генетических маркеров в селекционно-племенной работе со свиньями канадской селекции / Н.В. Проскурина [и др.] // Сельскохозяйственная биология. – 2007. — №6. – с. 41-47.
- Зиновьева Н.А. Генетическая характеристика свиней крупная белая и йоркшир различного происхождения с использованием ДНК-маркеров / Н.А. Зиновьева [и др.] // Российская сельскохозяйственная наука. – 2008. — №2. – с. 33-36.
- Зиновьева Н.А. Некоторые аспекты использования микросателлитов в свиноводстве / Н.А. Зиновьева, Е.И. Сизарева, Е.А. Гладырь // Достижения науки и техники АПК : ежемес. теорет. и науч.-практ. журн. — 2009. — № 8. — С. 38-41.
- Харзинова В.Р.. Локальные породы свиней: сравнительная характеристика аллелофонда на основе анализа микросателлитов / О.В. Харзинова Н.А, Костюнина, В.Р. Зиновьева // Свиноводство. – 2017. — №1. – с. 5-7.
- Инструкция по бонитировке северных оленей. Составители: НИИСХ Крайнего Севера, ВНИИ племенного дела с участием специалистов Управления Севера Госагропрома РСФСР, специалистов Госагропрома СССР, ЯНИИСХ, и др.- Н.: — 1988. — 19 с.
- Леханов Б.И. Домашнее оленеводство в горно-таежной зоне. — Спб.: НИИ географии СПБГУ. -2003. -108 с.