
Титульный лист и исполнители
РЕФЕРАТ
Отчет 102 с., 1 кн., 18 рис., 14 табл., 180 источн., 6 прил.
ОЗИМАЯ ПШЕНИЦА, ВРЕДИТЕЛИ, БИОИНСЕКТИЦИДЫ, ФИТОФАГИ, БИОЛОГИЧЕСКАЯ ЭФФЕКТИВНОСТЬ, УРОЖАЙНОСТЬ, ЭКОНОМИЧЕСКАЯ ЭФФЕКТИВНОСТЬ
Объектом исследования является сорт озимой пшеницы Юка.
Цель исследований – разработка системы биологической защиты озимой пшеницы в весенне-летний период вегетации от вредителей.
Схема опыта: контроль – без обработки; бионсектицид Биослип БВ – 3,0 л/га; Биослип БТ – 3,0 л/га; совместное применение препаратов Биослип БВ и Биослип ВТ – 1,5 + 1,5 л/га. Эталон смесь инсектицидов Алт-Альф, КЭ (альфа-циперметрин) норма расхода 0,1 л/га и Актара, ВДГ (тиаметоксам) норма расхода 0,05 кг/га.
В результате исследований впервые было установлено, что биоинсектицид Биослип БВ наиболее эффективен против злаковых тлей: биологическая эффективность 78,3-87,0 %. Действие биоинсектицида Биослип БТ против пьявицы красногрудой составляет 84,2%, что сопоставимо с химической защитой.
С точки зрения экономической эффективности обработка биоинсектицидом Биослип БВ наиболее рентабельна.
Результаты исследований можно рекомендовать для внедрения в технологию возделывания озимой пшеницы.
ВВЕДЕНИЕ
Приоритетной задачей России в научно-техническом развитии является переход к высокопродуктивному и экологически чистому агрохозяйству, разработка и внедрение систем рационального применения средств защиты сельскохозяйственных растений согласно «Стратегии научно-технологического развития Российской Федерации», утверждённой Указом Президента РФ (01.12.2016 года № 642).
Центральное Предкавказье, значительную площадь которого занимает Ставропольская возвышенность, по почвенно-климатическим условиям благоприятно для выращивания более 20 видов сельскохозяйственных растений, среди которых доминируют зерновые колосовые культуры.
Вместе с тем высокая концентрация посевов зерновых колосовых культур, особенно озимой пшеницы, наряду с рядом климатических и экологических факторов (мягкие непродолжительные зимы, высокая теплообеспеченность, длительный вегетационный период, наличие ряда культур — промежуточных хозяев вредных видов, широкая сеть лесополос — мест их резерваций, зимовки) создают благоприятные условия для развития и массового размножения многих видов вредителей, фитопатогенов и сорных растений.
Наблюдаемая дестабилизация фитосанитарного состояния посевов озимой пшеницы в Центральном Предкавказье свидетельствует о необходимости оптимизации зональной системы ее защиты от вредных видов применительно к ситуации, складывающейся с доминантными вредителями, способствующей стабильному функционированию ее агробиоценоза. Это требует новых подходов к решению проблемы.
В настоящее время наша страна по производству зерна пшеницы занимает лидирующие место в мире. Посевная площадь пшеницы составляет около 20 % всей мировой площади, занятой этой культурой. Основные площади посева озимой пшеницы размещены в районах с благоприятными условиями перезимовки – на Северном Кавказе, в Центрально – Черноземной зоне, а также в районах Поволжья.
Агробиоценозы озимой пшеницы в России представлены большим числом различных видов членистоногих, связанных между собой самыми разнообразными взаимоотношениями. При этом особое внимание следует уделять доминантным видам вредителей, которые дают регулярные вспышки массового размножения и причиняют значительный вред возделываемым сельскохозяйственным культурам. В России потери зерна от вредных организмов достигают 41,0 % при среднемировом уровне – 34%, потенциальные потери от вредителей составляют 13,1 %.
Одной из основных глобальных проблем современности является обострение экологической ситуации. В связи с этим экологизация сельскохозяйственного производства стала наиболее динамично развивающимся направлением аграрной науки XXI века, и нашла свое отражение в концепциях адаптивно-ландшафтного и органического земледелия, биологизированных системах защиты растений.
1 ОБОСНОВАНИЕ ВЫБОРА НАПРАВЛЕНИЯ ИССЛЕДОВАНИЙ
Наблюдаемое в настоящее время обострение экологической ситуации является одной из основных глобальных проблем современности, вызывая серьезную озабоченность мировой общественности. Развитие сельского хозяйства, особенно в период интенсивного вовлечения разнообразных ресурсов для повышения продуктивности культурных растений, оказывает все возрастающее воздействие на окружающую среду. Приводит к серьезным нарушениям в структуре видового состава членистоногих, фитопатогенов и сорных растений, появлению новых вредоносных видов вселенцев (инвазионных видов). В связи с этим наиболее динамично развивающимся направлением аграрной науки XXI века является экологизация сельскохозяйственного производства, нашедшее свое отражение в концепциях адаптивно-ландшафтного и органического земледелия, биологизированных и точечных систем и т.д. (Blackwell, 1994; Жученко, 2004; Kreue, et al., 1999; Dedryver et al., 2010; Новожилов, 2003; 2005; Шпаар, 2012; Павлюшин и др., 2013; Баздырев др., 2014 и др.).
Фундаментальным положением экологизации при выращивании сельскохозяйственных культур является использование для борьбы с популяциями вредных видов биологических агентов и препаратов, а также приемов, воздействующих на биоценотические связи в агробиоценозах (Мейер , 1937; Thompson, 1951; Суитмен, 1967; Бондаренко, 1986; Новожилов, 1993; Чертова, 1997; Bottcher, Gent, 2000; Smith, 2004; Aukema, 2005; Шпаар, 2008; Павлюшин и др., 2015).
Исследования энтомофауны агробиоценозов озимой пшеницы достаточно многочисленны, как в нашей стране, так и в других странах (Холодковский, 1902, 1914; Абдулхаирова, 1977; 1981; Dean, 1973; Grapel, Heinrich, 1980; Тряпицын, Шапиро, Щепетильникова, 1982; Марус, 2003; Harbaoui, et al., 2008; Frere, et al., 2011; Иванцова, 2009, 2014; Берим, 2014 и др.). При сравнении энтомофауны этой культуры в разных регионах мира (Европа, Азия, Северная и Южная Америки) обнаруживается значительное сходство её видового состава (Бусуек, 1977; Aukema, 2005). Вместе с тем прослеживается закономерная смена видов и их обилия, которая объясняется общностью связи местных энтомофаун с энтомофаунами других регионов (Шуровенков, 1977; Сливкин, 1983; Мутин,1985; Kokubu, Duelli, 1986; Carl, 1987; Кротова, 1989; Мансуров, Бабанов, Хамраев, 2002; Чекмарева, 2004; Tulin, 2011; Mawela etal., 2013).
В России потери зерна от вредных организмов достигают 41,0 % при среднемировом уровне — 34%, потенциальные потери от вредителей составляют 13,1 % (Шпаар, 2003, 2008, 2012; Olanca et al., 2009).
Агробиоценозы озимой пшеницы в России представлены большим числом различных видов членистоногих, связанных между собой самыми разнообразными взаимоотношениями (Воронин и др., 1986; Поляков, Копанева, 1986; Ченикалова и др., 2008; Белицкая, Иванцова, 2012). Это предусматривает необходимость постоянного мониторинга фитосанитарного состояния посевов озимой пшеницы с учётом взаимоотношений между растением и фитофагами (Поляков, 1983; Basky, Hopper, 2000; Нейморовец, 2008; Гриценко, 2010; Boivin et al., 2012; Шушковская, 2013; Захаренко, 2014). При этом особое внимание следует уделять доминантным видам вредителей, которые дают регулярные вспышки массового размножения и причиняют значительный вред возделываемым сельскохозяйственным культурам. (Павлюшин и др., 2015).
Такие виды должны держаться постоянно в поле зрения специалистов, так как их видовой состав и причиняемый вред посевам озимой пшеницы варьируют по регионам товарного производства зерна и в значительной степени определяются различными экзогенными факторами (Викторов, 1967; Бигон, 1989; Зазимко, 1995; Степановских, 2001; Sigsgaard, 2002; Beckage, Gelman, 2004; Islamoglu et al., 2010; Mawela et al., 2013; Шпанев и др., 2017).
Экзогенные факторы подразделяют на абиотические, биотические и антропогенные. Абиотические факторы, в свою очередь, делятся на химические (газовые составляющие атмосферного воздуха, химический состав почв и т.д.), климатические (температура, влажность, интенсивность солнечного излучения и т.д.) и эдафические (почвенные), в отдельную группу выделяют орографические (рельеф, геологические различия земной поверхности и т.д ) (Поляков, 1968; 1976; Чернышев, 1996). К биотическим факторам относятся трофические (пищевые), межвидовые и внутривидовые отношения (Шапиро, Вилкова, 1972, 1976; Шапиро, 1976; 1985; Шапиро и др., 1986). К антропогенным факторам относят различные воздействия человека на окружающую среду и экосистемы (распашка земель, полезащитное лесоразведение, агротехнические приёмы, применение пестицидов и т.д.) (Новожилов, 1997; Новожилов, Сухорученко, 1995; 1997; Жученко, 2004; 2010; Павлюшин и др., 2008; 2013).
Между перечисленными факторами существуют прямые и косвенные взаимосвязи, но действие каждого из них на функционирование агроэкосистем неоднозначно. Однако среди этих факторов можно выделить ряд ведущих, оказывающих существенное влияние на развитие и динамику численности вредных видов, их взаимоотношения с кормовым растением и энтомофагами.
Для успешной защиты озимой пшеницы от фитофагов необходим выбор способов и методов борьбы, и, прежде всего, выбор инсектицидов и их обоснованное и своевременное применение. Своевременность проводимых обработок способствует не только получению высокой эффективности защитных мероприятий для максимального получения количества и сохранения качества урожая, но для минимализации затрат на защиту пшеницы от комплекса вредителей. Но для этого нужен прогноз численности доминантных вредителей (Осмоловский, 1964).
О важности и необходимости прогноза фитофагов еще в начале 20 века было указано в трудах И. В. Курдюмова (1913). В 1929 г. был организован Всесоюзный научно-исследовательский институт защиты растений (ВИЗР), в состав которого вошел сектор учета и прогноза с 16 филиалами в главнейших сельскохозяйственных регионах и большим количеством опорных пунктов по СССР. Большой вклад в разработку прогнозов внесли энтомологи В. П. Поспелов, А. В. Знаменский, С. А. Предтеченский, А. А. Любищев, И. Д. Стрельников, И. А. Рубцов, А. С. Мончадский, Г. Я. Бей -Биенко, Я.И. Принц, В.Н. Щеголев, И.В. Кожанчиков, Г. К. Пятницкий, А. С. Данилевский, А. Ф.
Крышталь, О. И. Петруха, К.И. Ларченко, М. С. Гиляров, Б. В. Добровольский, В. П. Васильев (Поляков и др., 1984).
Исследованиями этих ученых было выявлено, что динамика численности организмов — сложный процесс, отражающий многообразную совокупность взаимоотношений организмов и среды. Развитие теоретических основ экологии и совершенствование методик полевого изучения популяций дает все больше доказательств тому, что колебания численности организмов — результат закономерного процесса, а не случайного сочетания различных факторов среды. В связи с этим концепция автоматической регуляции численности организмов в природе полуила широкое признание отечественных ученых (Сукачев, 1974; Посылаева, Сергиенко, 1996; Танский,1997; Соколов, 2001 и др.).
Признание факта колебания численности организмов как регулируемого процесса привело к необходимости изучения его механизмов. По степени воздействия на динамику популяций различные экологические факторы стали делить на группы: катастрофические и факультативные, нереактивные и реактивные, независимые и зависимые от плотности популяции, регулирующие и модифицирующие (Гиляров, 1955; Викторов, 1971; Одум, 1975; Carlson, 1988; Новожилов, 1991; Новожилов, Захаренко, 2000а; Попов, 2003; French et al., 2001; Gao, Liu, 2013).
Считается, что наблюдаемые в природе непрерывные изменения численности организмов — результат взаимодействия двух процессов: модификации и регуляции. Модификация обусловлена действием случайно меняющихся по отношению к рассматриваемой популяции факторов среды, главным образом, абиотического характера (Павлов, 1976; Feeney, 1987; Васильев, 1988; Mayers et al., 1989 Minaei, Mound, 2008). Однако при наблюдениях в природных условиях довольно сложно понять, какие из факторов среды наиболее существенно влияют на численность популяции (Киль, др., 2007).
История изучения динамики численности вредителей озимой пшеницы характеризуется последовательным проведением исследований по установлению значения отдельных факторов этой динамики, рассмотренных выше. В связи с несовершенством и трудоёмкостью методических разработок, оценка интенсивности воздействия этих факторов проводилась по относительной шкале или отдельно от других факторов. При этом комплексного охвата основных факторов динамики численности фитофагов и их энтомофагов не было.
Исключение составляют работы Г.А. Викторова (1967, 1976), в которых он анализирует одновременно несколько факторов, влияющих на динамику численности вредной черепашки: морфофункционального состояния клопов, погодных условий, фенологии кормового растения и вредителя, а так же активности природных энтомофагов.
В соответствии с такими представлениями о закономерностях динамики популяции фитофагов осуществлялась разработка методов прогноза их численности. Вначале эти методы основывались на основании одного или нескольких факторов среды без учета сложности их взаимосвязи между собой или с качественным и количественным состоянием популяции фитофага в течение всего периода его жизненного цикла.
Так, наблюдения в Польше за численностью пшеничного трипса (Haplothrips tritici Kurd), проведенные на протяжении 14 лет, показали четкую связь его численности с погодой (Luterek, 1981). Этот автор, используя метод множественной регрессии, получил следующее уравнение:
Log Y = — 2,390 + 0,125×1 + 0,202×2 + 0,186х3 + 0,085×4, где Y — реальная численность популяции; х1 — сумма эффективных температур за сезон;
х2 — суммарное количество осадков за два первых весенних месяца; х3 — средняя эффективная температура за эти же месяцы; х4 — значение X1 за предыдущий год.
Учитывая данные факторы, исследователь смог объяснить до 84% колебаний численности этого вида.
Однако этот пример, показывающий статистическую связь численности популяций данного вида трипса с погодными факторами, все же не доказывает, что только погода влияет на численность насекомых.
Исследования И.Я. Полякова, Л.А. Макаровой и Г.М. Дорониной (1985) установили, что популяции вредной черепашки из различных регионов страны различаются по специфичности реакции на абиотические факторы. Были выявлены погодные показатели развития клопа в основных районах его обитания и рассчитаны аналитические уравнения регрессии для годичного прогноза численности этого фитофага. Так ожидаемая численность для будущего года рассчитывалась по следующим формулам: для южных районов ареала: у = 0,081х1 — 0,512х2 + 0,04х 3 — 0,075 ± 0,156; для восточных районов: у = 0,064х1 + 0,061х3 — 0,077х4 — 0,472 ± 0,120, где у — уровень численности в будущем году; х1 — средняя температура периода лёта перезимовавших клопов; х2 — ГТК периода массовой яйцекладки и отрождения личинок; х3 — средняя температура периода развития личинок; х4 — количество «критических» декад за зиму прогнозируемого периода.
Было установлено, что при значении уравнения регрессии менее 1,4 следует ожидать снижения численности вредителя, более 1,6 — ее нарастания по сравнению с текущим годом. Анализ фактических и расчетных уровней численности в Ростовской области за 42 года показал, что оправдываемость такого прогноза составляет не менее 80%.
От погоды, несомненно, зависит состояние и кормового растения, а также активность паразитов и патогенных микроорганизмов (Tanzubil, 2000; Чернышев, 2006; Бокина, 2009). Погода оказывает первичное воздействие на жизненную систему, выводя ее из равновесного состояния, приводя прямо или косвенно либо к гибели значительной части популяции, либо способствуя ее расцвету (Косов, Поляков, 1958).
В дальнейшем по мере углубления изучения биологии и экологии фитофагов прогнозы стали дополняться новыми критериями, отражающими прямое или косвенное воздействия внешний условий на уровень и состояние популяции вредителей. С 70-х гг. прошлого столетия и по настоящее время прогнозом численности доминантных видов фитофагов различных культур ежегодно занимаются республиканские, краевые и областные ФГБУ «Россельхозцентры», на федеральном уровне — Российская лаборатория диагностики и прогнозов. Эти организации разрабатывают долгосрочный прогноз (на ближайший год) развития наиболее вредоносных фитофагов, используя показатели об обследованных и заселенных ими площадях, средней и максимальной численности, объемах обработок, фенологии, плодовитости, выживаемости и т.д. (Фролов, 2011). При этом прогнозы не приводят конкретных цифр численности вредителей, прогнозируемое ее изменение выражается в тенденции этого изменения (вероятен спад ли подъем численности) (Стамо. 2017 и др.).
Наиболее важным для сельхозяйственных товаропроизводителей является краткосрочный прогноз. Он позволяет заранее предвидеть время появления и продолжительность развития каждой стадии развития насекомого. Долгое время для прогнозирования использовали показатель, «сумма эффективных температур» (СЭТ). Н.М. Виноградовой (1970) было установлено, что для развития яиц вредной черепашки требуется, чтобы СЭТ отсчитывали выше 10° и равнялась она 95°, для личинок I возраста — 55°, 11- го возраста — 90° и III возраста — 65°. Раньше сигналом для начала инсектицидных обработок было значение «суммы эффективных температур» равное 240-280° от начала массовой откладки яиц клопами (Бойко, 1984).
А.С. Подольским (1974) был разработан метод «суммы индексов развития за сутки с помощью температурно-фенологических номограмм». Данный метод выражается в процентах и позволяет определить сроки развития любой стадии развития вредителя с точностью до 1 -2 дней и своевременно сигнализировать о начале применения инсектицидов.
Но краткосрочный прогноз так же не отражает показатели численности популяции того или иного объекта в определенном агробиоценозе или хозяйстве. В связи с этим необходимо отметить, что во-первых, современная система защиты озимой пшеницы нуждается в четких критериях прогноза, на основании которых можно было бы судить не только о тенденции изменения численности вредителей, но и о конкретных показателях их численности. Во-вторых, при использовании математического анализа агроклиматических критериев прогнозов учитывается лишь аспект воздействия абиотических факторов на динамику численности фитофагов. Между тем, известно, что воздействие этих факторов часто преломляются через систему трофических и биоценотических отношений фитофагов и энтомофагов с наложением на них технологических приемов возделывания озимой пшеницы (Шапиро и др. 1979).
Таким образом, многообразие и сложность существующей системы факторов прямого и косвенного воздействия на основные показатели популяционной изменчивости вредных видов в агробиоценозах озимой пшеницы свидетельствуют о том, прогнозирование их численности и вредоносности возможно только на основании использования комплекса математических моделей и разработки на их основе компьютерных программ. Для этого необходимо получение целого массива данных по изменению численности доминантных видов вредителей с учетом действия на них энтомофагов на разных этапах онтогенеза озимой пшеницы под влиянием меняющихся погодных условий, основных приемов технологии ее выращивания и отдельных экологических элементов.
Аналогичного мнения придерживается А.Н. Фролов, который в своей фундаментальной статье (2017) пишет «Математическое моделирование динамики численности остается одной из центральных проблем математической экологии и наиболее перспективны нелинейные модели, важная задача которых выявить точки неустойчивости системы, как эффективные состояния с точки зрения теории управления».
Считается, что при решении вопросов организации защиты растений в в крупных агроэкосистемах, необходимо осознавать, что борьба с вредными видами может быть достигнута только при определённых уровнях их развития и допустимых потерях продукции (Захаренко, Судариков, 2000). На зерновых культурах эти потери от вредителей можно довести до 3-4 %. Допуская такие потери и используя различные приёмы защитных мероприятий, по мнению этих авторов, можно добиться стабилизации численности вредных видов на приемлемо низком уровне.
Фитосанитарное состояние посевов озимой пшеницы за последние годы заметно ухудшилось. К этому приводит сев пшеницы по стерневым предшественникам, наличие падалицы, сорняков, сев в ранние или же слишком поздние сроки, завышение норм высева семян, посев по свежевспаханной почве, а также по минимальной обработке, и игнорирование других агротехнических приемов.
Произошедшие в 90 гг. прошлого столетия изменения в социально- экономических и производственных отношениях в аграрном комплексе России привели к резкому ухудшению фитосанитарного состояния посевов многих сельскохозяйственных культур, в том числе и озимой пшеницы в ряде регионов, включая Центральное Предкавказье.
Наблюдаемые в Ставропольском крае изменения в структуре землепользования, появление хозяйств разных форм собственности, возникшие финансово-экономические трудности у сельхозпроизводителей негативно сказались на функционировании зональной системы возделывания озимой пшеницы. Так произошедшее в 1995-2015 гг. насыщение структуры посевных площадей озимой пшеницей (более 58 %) на фоне их сокращения с 3433,9 до 2624,6 тыс. га, переход на укороченные севообороты (5-6-польные) и поверхностную обработку почвы на глубину 8-12 см, уменьшение в 7-10 раз объемов использования минеральных и органических удобрений привело к снижению плодородия, биологической активности и супрессивности почвы (Дорожко и др., 2011; Коваленков, 2000; Коваленков, Тюрина, 2016). Следствием этого явилось усиление развития и распространения корневых гнилей разной этиологии (Fusarium spp., Cercosporella herpotrichoides (Fron) Deighton, Gibellina cerealis Pass и др.) и аэрогенной инфекции, представленной мучнистой росой (Erysiphe graminis f. tritici), бурой ржавчиной (Puccinia triticina Erikss. ), септориозом (Septoria tritici var. lolicola R. Sprague & Aar.G. Johnson и S. nodorum Berk. & M.A. Curtis), несмотря на достаточно большие объемы применения фунгицидов (Шутко и соавт., 2011, Шутко, 2013). Наблюдается накопление в почве семян и в посевах озимой пшеницы озимых и зимующих сорных растений подмаренника цепкого (Galium aparine L.), василька синего (Centaurea cyanus L.), дескурении Софьи (Descurainia Sophia L.), ромашки непахучей (Matricaria perforate Merat.), костера кровельного — (Anisantha tectorum (L.) Nevski) и др. (Боташева, 2002, 2002а).
Ситуация с вредителями озимой пшеницы в Ставропольском крае на протяжении нескольких десятков лет существенно менялась. В прошлом столетии в северо-западной зоне края значительный вред посевам причиняла шведская муха (Oscinella frit L.), хлебная жужелица (Zabrus tenebrioides Goeze), наблюдались спорадические вспышки размножения красногрудой пьявицы (Lema melanopus L.), в 2001 г. распространилась и стала повреждать посевы пшеничная муха (Fhorbia fumigate Meigen).
Однако внедрение интенсивной технологии возделывания озимой пшеницы, направленной на получение высоких урожаев за счет резкого увеличения объемов использования средств химизации, включая пестициды, позволяло эффективно бороться с комплексом этих вредителей и снизить их численность ниже экономически опасного уровня. Единственным вредителем, против которого продолжали проводить регулярные химические обработки посевов был клоп вредная черепашка (Erygaster integriceps Put.), численность и вредоносность которого оставались высокими («Россельхозцентра» по Ставропольскому краю «Прогноз фитосанитарного состояния посевов … на 2015 г.).
В начале 2000 гг. ситуация и с вредителями озимой пшеницы значительно осложнилась. На культуре сформировался комплекс сосущих вредителей, состоящий из нескольких доминантных видов, ежегодно развивающихся на ее посевах в периоды роста вегетативной массы и формирования репродуктивной сферы. Этот комплекс представлен вредной черепашкой, сохранившей статус одного из наиболее опасных вредителей озимой пшеницы на Ставрополье и в XXI веке, а также пшеничным трипсом (Haplothrips tritici Kurd.), большой и обыкновенной злаковыми тлями (Sitobion avenae F., Schizaphis graminum Rond.), обыкновенным и черным хлебными пилильщиками (Cephus pygmaeus L., Trachelus tabidus F.), не имевших ранее экономического значения (Алексеев, 2003; Скребцова, 2009; Щербакова, 2009; Мухина, Шабалдас, Солодилина, 2013; Коваленков, 2014).
В настоящее время происходит не только увеличение численности и вредоносности этих фитофагов, но и расширение видовых ареалов вредной черепашки, хлебных пилильщиков и пшеничного трипса (Алексеев, Демкин, 2005; Скребцова, 2009; Пикушова и др., 2011). Росту численности этих вредителей на озимой пшенице способствует также длительное и бесконтрольное применение на больших площадях фосфорорганических препаратов и пиретроидов, эффективность которых резко снизилась (до 45,5-60%) вследствие развития к ним резистентности (Коваленков, Тюрина, 2016).
Несмотря на то, что обработки посевов озимой пшеницы против указанных вредителей инсектицидами доходят до 1550 тыс. га («Россельхозцентра» по Ставропольскому краю «Прогноз фитосанитарного состояния посевов … на 2017 г.), потери урожая зерна от их общей деятельности по нашим подсчетам достигают в среднем 5-7 ц/га ежегодно (Глазунова, 2014) при значительном повреждении зерна вредной черепашкой. Например, по данным специалистов ФГБУ «Россельхозцентр» в 2011 г. в Ставропольском крае вредной черепашкой было повреждено 2131 тыс. тонн зерна озимой пшеницы при проценте поврежденности от 0,6 до 12% (Стамо, Кузнецова, 2012).
У клопа вредной черепашки, пшеничного трипса, хлебных пилильщиков и злаковых тлей за последние двадцать лет в Ставропольском крае отсутствуют спады и подъемы численности, их развитие приобрело характер ускоренного нарастания численности и расширение видовых ареалов. Они стали доминантными компонентами агробиоценоза озимой пшеницы. Благодаря широкому спектру адаптаций в их популяциях стало происходить нарастание по годам показателей развития резистентности к инсектицидам с последующим спадом эффективности применяемых препаратов.
Одной из основных глобальных проблем современности является обострение экологической ситуации. В связи с этим экологизация сельскохозяйственного производства стала наиболее динамично развивающимся направлением аграрной науки XXI века, и нашла свое отражение в концепциях адаптивно-ландшафтного и органического земледелия, биологизированных системах защиты растений. Приоритетной задачей России в научно-техническом развитии является переход к высокопродуктивному и экологически чистому агрохозяйству, разработка и внедрение систем рационального применения средств защиты сельскохозяйственных растений согласно «Стратегии научно-технологического развития Российской Федерации», утверждённой Указом Президента РФ (01.12.2016 года № 642).
В настоящее время биологические приемы защиты растений находят все более широкое применение, расширяется арсенал средств биометода.
В странах СНГ в более чем 500 республиканских и районных (краевых, областных) биолабораториях было налажено производство трихограммы, энкарзии (паразита белокрылки), хищной галлицы афидимизы, клеща фитосейулюса, а также биопрепаратов – боверина, БИП, битоксибациллин, дробацилина, бактороденцида и другие.
На территории РФ впервые в мировой практике были созданы крупные биофабрики по производству трихограммы, разработаны автоматизированные линии по разведению златоглазок, получению трихограммы в желатиновых капсулах с ИПС вместо яиц хозяина.
В страну было завезено более 25 видов энтомофагов и микроорганизмов для борьбы с вредителями.
В последние годы получили широкое развитие принципиально новые направления в защите растений: использование в борьбе с вредными насекомыми гормонов и их синтетических аналогов, половых аттрактантов, методов стерилизации насекомых, которые оцениваются как весьма перспективные и безопасные для человека, полезных животных и биосферы в целом.
Вещества, выделяемые железами внутренней секреции и регулирующие жизненные процессы живых организмов, называют гормонами. Этот термин впервые ввел английский физиолог Э. Старлинг ещё в 1905 г. Так, недостаток ювенального гормона развития насекомых вызывает появление особей с недоразвитыми крыльями, половой системой и другими изменениями, а его избыток замедляет развитие насекомых, вызывая появление добавочных возрастов личинок, задерживая окукливание.
Химическая стерилизация насекомых, например, выпуск стерильных самцов приводит также к отсутствию потомства вредителя. Этот метод предложил советский ученый А.С.Серебровский и успешно применял против комнатных мух, амбарного долгоносика и других. В дальнейшем метод успешно применен против мясной мухи на островах Тихого океана американскими энтомологами и против средиземноморской плодовой мухи в Греции. Метод успешнее применим на островных территориях из-за непоступления на территорию новых фертильных особей вредителей.
Использование синтетических (половых аттрактантов, феромонов) уже находит широкое применение для борьбы и сигнализации появления вредителей, например листоверток.
Идентифицированы и феромоны более чем 670 видов других вредителей. В США, Германии, Японии, Англии и других странах производством феромонных ловушек занято более 10 фирм, выпускающие ловушки для 30 и более видов вредителей сельского и лесного хозяйства.
Но каким бы эффективным ни был тот или иной метод борьбы с вредителями болезнями или сорняками, он не может решить проблему полностью. Поэтому будущее защиты растений за интеграцией, объединением всех методов в единую систему интегрированной защиты растений.
Идея интегрированной системы защиты растений зародилась еще в 30-е годы, когда видные ученые Н.В. Курдюмов, В.Н. Щеголев, Г.Я. Бей-Биенко, А.В. Знаменский и другие сформулировали ее основные принципы. В современном понимании интегрированная защита растений – это борьба с вредными организмами, учитывающая пороги их вредоносности и использующая в первую очередь природные ограничивающие факторы наряду с применением всех других методов, удовлетворяющих экономическим, экологическим и токсикологическим требованиям.
Интегрированная защита растений предусматривает выбор таких средств подавления вредных видов, которые не только сохраняли, но и активизировали бы деятельность полезных организмов.
Это идеальная комбинация биологических, агротехнических, химических, физических и других методов защиты растений против комплекса вредителей и болезней в конкретной эколого-географической зоне на определенной культуре.
Активизация научных исследований по разработке принципов интегрированных систем защиты растений и конкурентных их схем в ряде стран мира позволила достичь определенных результатов на хлопчатнике, плодовых, сахарной свекле, сое, табаке, овощных и других культурах.
Основными элементами интегрированных систем защиты растений являются:
достоверный централизованный и районный прогноз распространения, развития и численности вредных организмов;
учеты численности вредителей и развития болезней на посевах в хозяйстве с опорой на экономические пороги вредности объектов;
переход от сплошных химических обработок к выборочным, правильный выбор сроков и норм их применения;
широкое применение биологических средств защиты растений от вредителей и болезней.
правильная система агротехники и семеноводства, применение устойчивых сортов.
В связи со сложившейся ситуацией возникает необходимость в оптимизации системы защиты озимой пшеницы и поиску альтернативным решениям, по борьбе с доминантными видами вредителей в Центральном Предкавказье.
Целью исследования является проведение научных исследований по разработке системы биологической защиты озимой пшеницы в весенне-летний период вегетации против комплекса вредителей в природно-климатических условиях Ставропольского края.
В задачи наших исследований входило:
– выявление структуры и особенностей функционирования системы триотрофа – основной средообразующей консорции агробиоценоза на разных этапах органогенеза озимой пшеницы;
– изучение биологических особенностей развития фитофагов, наносящих вред в период формирования репродуктивных органов озимой пшеницы;
– определение биологической эффективности биоинсектицидов для различных групп фитофагов озимой пшеницы;
– определение токсичности биоинсектицидов для различных групп энтомофагов вредителей озимой пшеницы;
– расчёт экономической эффективности биологической защиты озимой пшеницы от вредителей;
– выводы и рекомендации на основе полученных результатов
2. УСЛОВИЯ ПРОВЕДЕНИЯ ИССЛЕДОВАНИЙ
2.1. Почвенно-климатические условия места исследований
Исследования проводились в 2019 году на территории учебно-опытной станции ФГОУ ВО «Ставропольский государственный аграрный университет». Она находится на территории Шпаковского района Ставропольского края, юго-восточнее г. Ставрополя.
Учебно-опытное хозяйство организовано в 1932 году. В его состав входят: поселок Демино, хутора Холодногорский и Гремучий. Расположено на северном склоне Ставропольской возвышенности в Грачевско-Калаусском ландшафте лесов и степей и на юго-западном склоне Верхнеегорлыкского водораздельного ландшафта типичных лесостепей, имеет сложную конфигурацию. Землепользование учхоза сильно расчленено. Более ¾ сельскохозяйственных угодий расположено на склонах крутизной более 1о, что способствует развитию эрозионных процессов. Большое количество балок чередуются с довольно высокими увалами, имеющими в ряде мест крутые склоны с выходом на поверхность материнских пород.
Территория учебно-опытной станции (учхоз) расположена в III агроклиматической зоне Ставропольского края. Климатические условия обусловлены влиянием вертикальной зональности (высота над уровнем моря 500-550 м) и резко-континентальным климатом прилегающих районов.
Характерной особенностью зоны является неустойчивое увлажнение по годам и неравномерность выпадения осадков в течение года. Средняя многолетняя сумма осадков составляет 623 мм, за вегетационный период выпадает 350-370 мм. Гидротермический коэффициент 1,2-1,5. За вегетационный период. Сумма температур за период активной вегетации колеблется от 2800 до 3000˚С. Нарастание температур идет быстро. Средняя многолетняя температура самого теплого месяца – июля +23,9˚С, средняя месячная температура холодного месяца – января составила –3,7˚С. Максимальная температура в зимнее время опускается до -32˚С. Продолжительность зимы колеблется от 95 до 110 дней. Почва промерзает на 25-30 см, а в отдельные годы до 100 см. Снежный покров неустойчив, его средняя высота составляет 15-20 см.
Зима неустойчивая, длится 85-110 дней. Максимальная глубина промерзания почвы 27-29 см, а в отдельные годы до 100 см. Снежный покров неустойчив, средняя высота его 10-12 см. В течении зимы очень часто оттепели.
В зимний период преобладают восточные ветра. Весенние заморозки заканчиваются в апреле, иногда отмечаются и в мае. Среднесуточная температура воздуха поднимается выше +10˚С после 15-20 апреля. Перепад суточных температур через отметку + 5˚С происходит, как правило, весной в начале апреля и осенью во второй декаде ноября. Лето довольно жаркое, максимальная температура может достигать + 40˚С.
Высокие температуры обуславливают большую испаряемость, которая превышает количество выпадающих осадков.
Относительная влажность воздуха характеризует степень насыщенности воздуха водяными парами, которые оказывают большое влияние на развитие растений. В июле-августе относительная влажность воздуха опускается до 59-62%, что оказывает неблагоприятное действие на развитие растений.
Не редким явлением на территории опытной станции являются засухи и суховеи. Общее число дней с суховеями достигает 50-60. Суховеи могут сопровождаться сильными ветрами со скоростью более 15 м/с. накопление влаги в почве осуществляется преимущественно за счет осадков холодного периода, чему способствует неглубокое промерзание почвы, частые оттепели и невысокое испарение зимой.
Среднегодовая температура воздуха равна +7,5˚С, а почвы +9,3…+10˚С.
Высокие температуры тёплого периода обуславливают большую величину испаряемости, превышающую количество выпадающих осадков.
Относительная влажность воздуха характеризует степень насыщенности воздуха водяными парами, которые оказывают большое влияние на развитие растений. В июле — августе относительная влажность воздуха опускается до 65-59%, что оказывает неблагоприятное действие на развитие растений
Накопление влаги в почве осуществляется преимущественно за счёт осадков холодного периода, чему способствует неглубокое промерзание почвы, частые оттепели и невысокое испарение зимой (таблица 1, 2).
К положительным сторонам климата относятся длительный вегетационный период и высокая сумма положительных температур; к отрицательным – ливневый характер осадков и их неравномерное распределение по временам года, частые оттепели и, как следствие этого крайне неустойчивый снежный покров, суховеи.
Таблица 1 – Основные агроклиматические показатели по данным метеостанции г Ставрополя
| Показатели | Величины |
| Среднегодовая температура воздуха (°C) | 9,2 |
| Сумма температур за период с t≥+10°C (°C) | 2800-3200 |
| Годовая сумма осадков (мм) | 550-650 |
| в т.ч. за период с t≥10°C (мм) | 350-400 |
| Гидротермический коэффициент | 1,1-1,3 |
| Запасы продуктивной влаги к началу вегетации в слое почвы 0-100 см (мм) | 160-200 |
| Число суховейных дней | 61 |
| Продолжительность безморозного периода (дней) | 180-190 |
В общем можно сказать, что климатические условия, в основном, благоприятные для возделывания большинства сельскохозяйственных культур.
Учебно-опытная станция расположена на высоко эрозионноопасной глубокорасчлененной равнине. Высокие отметки и степень расчлененности поверхности убывает с юга на север. Пологие и слабопокатые склоны крутизной до 5º благоприятны для механизированной обработки, ухода за посевами, посева, уборки урожая и отведены под пашню.
Особенностью рельефа местности является слабая дренированность территории, наличие водонепроницаемых пород, верховодность, что привело к значительному увеличению площади подтопленных земель и образованию оползневых явлений.
В целом рельеф местности учебно-опытной станции позволяет организовывать ведение земледелия на высоком агротехническом уровне.
Землепользование учебно-опытного хозяйства расположено в зоне ковыльно-типчаково-разнотравяной степи. Целинная растительность при интенсивном ведении хозяйства сохранилось лишь на участках непригодных к распашке.
Почвенный покров хозяйства довольно однороден и почвы залегают здесь большими контурами. Почвы хозяйства представлены черноземом выщелоченным, тяжелосуглинистым.
Таблица 2 — Средние многолетние температура и осадки по данным метеостанции г. Ставрополь.
| Декада | Месяцы | Сумма за год | |||||||||||
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | ||
| Осадки, мм | |||||||||||||
| 1 | 12 | 9 | 10 | 15 | 22 | 29 | 29 | 20 | 19 | 15 | 15 | 14 | 623 |
| 2 | 10 | 9 | 11 | 18 | 23 | 31 | 27 | 17 | 18 | 14 | 16 | 14 | |
| 3 | 10 | 9 | 13 | 20 | 25 | 30 | 24 | 16 | 17 | 14 | 15 | 13 | |
| Температура, ºС | Сумма активн. температур | ||||||||||||
| 1 | -3,2 | -3,8 | -0,6 | 6,5 | 13,4 | 18,0 | 21,1 | 21,8 | 17,8 | 12,0 | 5,3 | 0,1 | 2800 |
| 2 | -3,8 | -3,0 | 1,6 | 8,5 | 15,2 | 19,0 | 22,0 | 21,6 | 16,0 | 10,0 | 3,4 | -1,1 | |
| 3 | -4,1 | -2,2 | 3,1 | 10,2 | 16,9 | 20,1 | 20,7 | 20,0 | 14,2 | 8,1 | 1,6 | -2,3 | |
Содержание гумуса в пахотном слое варьирует от 5,8 до 6,2%. Запасы гумуса в метровом слое достигают 500-550 т/га. Содержание подвижного фосфора по Мачигину – 22-26 мг, обменного калия – 290 мг/кг почвы. Характеристика почв приведена в таблице 3.
Таблица 3 — Общая характеристика почв учебно-опытного хозяйства
| Наименование почв | Глубина слоя, см | Общая масса, г/см3 | Удельная масса, г/см3 | рН водной вытяжки | Пахотный слой, см | Наименьшая влагоемкость % и сухой массы | Содержание гумуса | Содержание в 1 кг | |
| Р2О5 | К2О | ||||||||
| Чернозем выщелоченный тяжелосуглинистый | 0-40 | 1,20 | 2,55 | 6,7 | До 35 | 33,17 | 6,7 | 0,18 | 2,4 |
| 40-50 | 1,22 | 2,57 | 32,30 | ||||||
| 50-60 | 1,25 | 2,60 | 31,30 | ||||||
| 60-80 | 1,27 | 2,64 | 30,90 | ||||||
| 80-160 | 1,30 | 2,66 | 29,74 | ||||||
| Чернозем слобовыщелоченный слабосуглинистый | 0-40 | 1,22 | 2,56 | 6,5 | До 35 | 32,25 | 6,4 | 0,20 | 3,0 |
| 40-50 | 1,25 | 2,58 | 31,40 | ||||||
| 50-60 | 1,29 | 2,60 | 30,37 | ||||||
| 60-80 | 1,21 | 2,61 | 29,50 | ||||||
| 80-160 | 1,33 | 2,65 | 28,22 | ||||||
| Чернозем обыкновенный карбонатный тяжелосуглинистый | 0-40 | 1,26 | 2,58 | 7,1 | До 35 | 30,07 | 6,4 | 0,19 | 2,9 |
| 40-50 | 1,28 | 2,60 | 29,45 | ||||||
| 50-60 | 1,31 | 2,62 | 29,32 | ||||||
| 60-80 | 1,34 | 2,64 | 28,80 | ||||||
| 80-160 | 1,36 | 2,65 | 28,07 | ||||||
Почвы отличаются высокой емкостью поглощения, обусловленной высоким содержанием высокодисперсных илистых частиц. Емкость поглощения пахотного слоя 40 мг. экв/100 г. почвы.
Почвы имеют довольно плотное сложение 1,15-1,36 г/см3. реакция почвенного раствора слабокислая и нейтральная рН 5,6-6,5. почвы имеют среднюю обеспеченность подвижным фосфором, среднюю и высокую по калию.
Почвы учебно-опытного хозяйства обладают высоким плодородием, имеют хорошую зернисто-комковатую структуру (горизонт А), высокую гумусированность, оптимальную реакцию почвенного раствора, достаточное содержание основных элементов питания, отсутствие вредных солей, а также удачно сочетаются здесь с благоприятными климатическими условиями, что позволяет ежегодно получать способные и высокие урожаи сельскохозяйственных культур.
2.2. Метеорологические условия в период проведения опытов
По среднегодовым показателям вегетационный год 2018-2019 произрастания озимой пшеницы сильно отличался от среднемноголетних значений. Год был неблагоприятен для озимых культур.
Сентябрь 2018 г. Норма среднемесячной температуры сентября: 16.4°. Фактическая температура месяца по данным наблюдений: 18.1°. Отклонение от нормы: +1.7°. Самая низкая температура воздуха (5.2°) была 28 сентября. Самая высокая температура воздуха (29.7°) была 2 сентября. Норма суммы осадков в сентябре: 47 мм. Выпало осадков: 42 мм. Эта сумма составляет 89% от нормы.
Октябрь 2018 г. Норма среднемесячной температуры октября: 10.0°. Фактическая температура месяца по данным наблюдений: 12.4°. Отклонение от нормы: +2.4°. Самая низкая температура воздуха (0.0°) была 6 октября. Самая высокая температура воздуха (24.8°) была 1 октября. Норма суммы осадков в октябре: 49 мм. Выпало осадков: 42 мм. Эта сумма составляет 85% от нормы.
Ноябрь 2018 г. Норма среднемесячной температуры ноября: 3.4°. Фактическая температура месяца по данным наблюдений: 2.0°. Отклонение от нормы: -1.4°. Самая низкая температура воздуха (-8.7°) была 24 ноября. Самая высокая температура воздуха (16.8°) была 4 ноября. Норма суммы осадков в ноябре: 46 мм. Выпало осадков: 48 мм. Эта сумма составляет 105% от нормы.
Декабрь 2018 г. Норма среднемесячной температуры декабря: -0.7°. Фактическая температура месяца по данным наблюдений: -0.0°. Отклонение от нормы: +0.7°. Самая низкая температура воздуха (-6.8°) была 17 декабря. Самая высокая температура воздуха (7.7°) была 6 декабря. Норма суммы осадков в декабре: 33 мм. Выпало осадков: 45 мм. Эта сумма составляет 135% от нормы.
Январь 2019 г. Норма среднемесячной температуры января: -2.3°. Фактическая температура месяца по данным наблюдений: -0.5°. Отклонение от нормы: +1.8°. Самая низкая температура воздуха (-9.0°) была 10 января. Самая высокая температура воздуха (10.2°) была 16 января. Норма суммы осадков в январе: 29 мм. Выпало осадков: 24 мм. Эта сумма составляет 83% от нормы.
Февраль 2019 г. Норма среднемесячной температуры февраля: -2.3°. Фактическая температура месяца по данным наблюдений: 0.3°. Отклонение от нормы: +2.6°. Самая низкая температура воздуха (-8.2°) была 20 февраля. Самая высокая температура воздуха (8.8°) была 21 февраля. Норма суммы осадков в феврале: 28 мм. Выпало осадков: 20 мм. Эта сумма составляет 71% от нормы.
Март 2019 г. Норма среднемесячной температуры марта: 2.3°. Фактическая температура месяца по данным наблюдений: 4.0°. Отклонение от нормы: +1.7°. Самая низкая температура воздуха (-5.8°) была 4 марта. Самая высокая температура воздуха (18.8°) была 10 марта. Норма суммы осадков в марте: 35 мм. Выпало осадков: 45 мм. Эта сумма составляет 129% от нормы.
Апрель 2019 г. Норма среднемесячной температуры апреля: 9.6°. Фактическая температура месяца по данным наблюдений: 9.5°. Отклонение от нормы: -0.1°. Самая низкая температура воздуха (-2.3°) была 5 апреля. Самая высокая температура воздуха (23.6°) была 28 апреля. Норма суммы осадков в апреле: 45 мм. Выпало осадков: 22 мм. Эта сумма составляет 49% от нормы.
Май 2019 г. Норма среднемесячной температуры мая: 14.8°. Фактическая температура месяца по данным наблюдений: 17.1°. Отклонение от нормы: +2.3°. Самая низкая температура воздуха (6.4°) была 5 мая. Самая высокая температура воздуха (29.6°) была 31 мая. Норма суммы осадков в мае: 64 мм. Выпало осадков: 44 мм. Эта сумма составляет 68% от нормы.
Июнь 2019 г. Норма среднемесячной температуры июня: 19.2°. Фактическая температура месяца по данным наблюдений: 23.9°. Отклонение от нормы: +4.7° (РЕКОРД). Самая низкая температура воздуха (12.3°) была 30 июня. Самая высокая температура воздуха (34.9°) была 23 июня. Норма суммы осадков в июне: 83 мм. Выпало осадков: 28 мм. Эта сумма составляет 34% от нормы.
Июль 2019 г. Норма среднемесячной температуры июля: 22.3°. Фактическая температура месяца по данным наблюдений: 21.9°. Отклонение от нормы: +0.1°. Норма суммы осадков в июле: 58 мм. Выпало осадков: 23 мм. Эта сумма составляет 40% от нормы. Самая низкая температура воздуха (11.1°) была 10 июля. Самая высокая температура воздуха (33.5°) была 8 июля (http://www.pogodaiklimat.ru/monitor.php?id)
В целом данные погодные условия были неблагоприятны для развития озимой пшеницы. Из-за того, что с осени в ноябре резко наступили холода и пшеница не успела раскуститься и в зиму ушла в удовлетворительном состоянии. В дальнейшем затяжная и прохладная весна позволила растениям раскуститься, но суховей в конце мая и резкое повышение температур в июне с низкой влажностью воздуха (менее 50 %) привели к стрессу растений озимой пшеницы и резкому созреванию, что отразилось на выполненности зерна и привело к снижению урожайности культуры.
Закладка опыта по первому сроку внесения биоинсектицидов Биослип БТ, Биослип БВ, и их смесей на озимой пшеницы по определению их биологической эффективности и влиянию его на урожайность культуры на учебно-опытной станции СтГАУ проводили 21 мая 2019 года. Средняя температура воздуха составляла + 17,6 ºС. Ясно. Ветер был восточный – 3 м/с. Осадков в этот день не было. Погодные условия соответствовали регламентам применения пестицидов (http://www.pogodaiklimat.ru/monitor.php?id=34949&month=5&year=2019).

Рисунок 1 – Обработка опытов 21 мая 2019 г.
Закладка опыта по второму сроку внесения биоинсектицидов Биослип БТ, Биослип БВ, и их смесей на озимой пшенице по определению их биологической эффективности и влиянию на урожайность культуры в учебно-опытной станции СтГАУ проводили 28 мая 2019 года. Средняя температура воздуха составляла + 17,9 ºС. Малооблачно. Ветер был западный – 4 м/с. Осадков в этот день не было. Погодные условия соответствовали регламентам применения пестицидов (http://www.pogodaiklimat.ru/monitor.php?id=34949&month=5&year=2019).


Рисунок 2 – Обработка опытов 28 мая 2019 г.

Рисунок 3 – Обработка опытов 04 июня 2019 года 3-й срок внесения биоинсектицидов
Закладка опыта по третьему сроку внесения биоинсектицидов Биослип БТ, Биослип БВ, и их смесей на озимой пшеницы по определению его биологической эффективности и влиянию его на урожайность культуры на учебно-опытной станции СтГАУ проводили 4 июня 2019 года. Средняя температура воздуха составляла + 23,2 ºС. Ясно. Ветер был северо-восточный – 3 м/с. Осадков в этот день не было. Погодные условия соответствовали регламентам применения пестицидов (http://www.pogodaiklimat.ru/monitor.php?id=34949&month=6&year=2019)
3 МЕТОДИКА ПРОВЕДЕНИЯ ИССЛЕДОВАНИЙ И СХЕМЫ ОПЫТОВ
Исследования проводились в 2019 году в условиях учебно-опытного хозяйства ФГБОУ ВО Ставропольского ГАУ. В посевах озимой пшеницы сорта Юка, предшественник озимая пшеница.
Для разработки биологической системы защиты озимой пшеницы использовали бионсектициды производства фирмы BIONOVATIC: «Биослип БВ», Ж содержащего (Количество спор штамма Beauveria bassiana OPB-09): не менее 1×108 спор в 1 мл и «Биослип ВТ», П содержащего (жизнеспособные споры и термостабильный кристаллический эндотоксин штаммов B. thuringiensis): не менее 1·1011 КОЕ/г (кристаллов токсина в 1 г.) в сравнении с применением химических инсектицидов Алт-Альф, КЭ (альфа-циперметрин 100 г/л) и Актара, ВДГ (тиаметоксам 250 г/кг). Определяли биологическую эффективность бионсектицидов и инсектицидов в отношении пьявицы красногрудой, злаковых тлей, вредной черепашки, пшеничного трипса и хлебных пилильщиков в условиях зоны неустойчивого увлажнения Ставропольского края. Инсектициды применялись согласно схеме опыта таблица 3.
В опытах был контроль (без обработки) три варианта применения биоинсектицидов в трех кратном применении (21 мая в фазу колошения (BBCH 51-55); 28 мая в фазу цветения (BBCH 57-65); 4 июня в фазу молочной спелости (BBCH 65-69)) и эталон баковая смесь химических инсектицидов в однократном применении (28 мая в фазу цветения (BBCH 57-65)). Площадь одного варианта составляла 1 га. Повторность опыта трех кратная.
Объектами исследований были следующие виды фитофагов из отрядов: Coleoptera — Oulema melanopus L. (сем. Chrysomelidae); Hemiptera — Eurygaster integriceps Put., Eurygaster maura L., Eurygaster austriacus Schr., Eurygaster testudinaris L. (сем. Scutelleridae); Aelia acumunata L., Aelia rostrata L., Carpocoris fuscispinus Boh., Dolycoris baccarum L. (сем. Pentatomidae); Homoptera Sitobion avenae F., Schizaphis graminum Rond. (сем. Aphididae); Thysanoptera Haplothrips tritici Kurd. (сем.Phloeothripidae); Hymenoptera — Cephus pygmaeus L., Trachelus tabidus F (сем. Cephidae).
Таблица 4 – Схема опыта, нормы расхода препаратов
и сроки их применения в процессе вегетации озимой пшеницы
| № | Фаза развития озимой пшеницы | |||||
| Колошение (BBCH 51-55) | Цветение(BBCH 57-65) | Молочная спелость ( BBCH 65-69) | ||||
| Наименование препарата | Норма расхода препарата, л/га | Наименование препарата | Норма расхода препарата, л/га | Наименование препарата | Норма расхода препарата, л/га | |
| 1 | Контроль (без обработки) | — | — | — | — | — |
| 2 | Биослип БВ | 3,0 | Биослип БВ | 3,0 | Биослип БВ | 3,0 |
| 3 | Биослип ВТ | 3,0 | Биослип ВТ | 3,0 | Биослип ВТ | 3,0 |
| 4 | Биослип БВ +
Биослип ВТ |
1,5 + 1,5 | Биослип БВ +
Биослип ВТ |
1,5 + 1,5 | Биослип БВ +
Биослип ВТ |
1,5 + 1,5 |
| 5 | — | — | Алт-Альф, КЭ + Актара, ВДГ | 0,1 + 0,05 | — | — |
Система защиты озимой пшеницы:
Осенью семена были протравлены фунгицидом Мактим Форте с нормой расхода 1,5 л/т с добавлением иммуномодулятора Зеребро Агро с нормой расхода 0,15 л/т.
Весной в фазу «конец кущения начало трубкования» (Стадия BBCH 29-32) была проведена обработка гербициды + фунгицид + иммуномодулятор: гербицидами Бен Гур, ВДГ 0,02 кг/га в смеси с граминицидом Ягуар Супер, КЭ – 0,6 л/га + Триагро, КС – 1,0 л/га + Зеребро Агро – 0,1 л/га; в фазу «начало колошения» (Стадия BBCH 50-52) была проведена обработка фунгицид + иммуномодулятор: Страйк Форте, КС – 0,6 л/га Зеребро Агро — 0,1 л/га. Инсектициды были внесены согласно схеме опыта.
Метод внесения препаратов – наземная обработка самоходным опрыскивателем «ATLANTIQUE 3200» с нормой расхода рабочей жидкости 200 л/га согласно схеме опыта.
Учеты численности пьявицы красногрудой, клопов вредной черепашки, пшеничного трипса, хлебных пилильщиков и злаковых тлей в период вегетации в посевах озимой пшеницы были проведены до обработки и после обработки на 3 и 7 день, согласно методикам И.Я. Поляков и др., (1984) и В.П. Омелюта (1989).
Методика и техника проведения учетов будет зависеть от вида вредителей, особенностей их биологии и поведения. Подсчет клопов вредной черепашки и хлебных пилильщиков будет проводиться энтомологическим сачком по диагонали повторности будет браться проба из десяти взмахов сачка и таких учетов на каждой повторности будет 10 штук.
Учеты численности пьявицы будем проводит на 10 учетных площадках размером 0,25 м2 (0,5смх0,5см). В связи с неравномерным расселением насекомых на посевах и большим сосредоточением их по краям полей пробы будем брать на одинаковом расстоянии одну от другой. При этом их размещать в пределах одной повторности варианта в шахматном порядке. На учетных площадках тщательно будут осматриваться все растения озимой пшеницы и сорные растения, а также сухие стебли, комочки земли, на которых может откладывать яйца пьявица. Во время учетов на растениях каждой пробы подсчитывают раздельно по фазам развития количество обнаруженных яиц, личинок, куколок и взрослых насекомых.
Для учета численности злаковых тлей и пшеничного трипса будем использовать следующую методику. В конце трубкования — начале цветения отбирать в пробу главные стебли, в фазе молочная — полная спелость — отбирать колосья. В шахматном порядке на поле в 10 местах срезали по десять колосьев (стеблей) и складывать отдельно каждую пробу в полиэтиленовый мешочек с этикеткой. В лаборатории проводить разбор и анализ следующим образом. На бумагу, на которой начерчен круг, разделенный на 4 части, высыпать содержимое мешочка, тщательно вытряхивая вредителей из главных стеблей и колосьев, затем равномерно распределяя их по кругу, в одной четвертой будем подсчитывать, затем умножать на 4 и делили на 10. Таким образом, получать среднее количество вредителей на одном колосе (стебле) в пробе (Деров, 1986).
Биологическую эффективность инсектицидов будем определять по смертности вредителя учитывая численность вредителей во всех вариантах и проворностях до и на определенные дни после проведения опрыскивания. Затем будем вычислять процент смертности по каждой повторности, а также средний процент по каждому варианту. Биологическая эффективность будет определяться при сопоставлении с контролем по формуле:
С= С1 – С2 = 100*В — 100*в ,
А а
где: С — смертность вредителя с поправкой на контроль, %; С1 и С2 — соответственно смертность вредителя в варианте с обработкой и на контроле, %; А и а- соответственно общее число особей в обработанном и контрольном варианте, шт.; В и в — соответственно число погибших особей в обработанном варианте и на контроле, шт.
Биологическая урожайность озимой пшеницы будет определена в фазу полной спелости культуры. Учет урожая будет произведен поделяночно, путем прямого комбайнирования с последующим пересчетом на 14% влажность.
Экономическая эффективность применения инсектицидов будет рассчитана после уборки культуры по обще принятой методике (Нечаев, Парамонов, Бершицкий, 2016).
4 РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
4.1. Структура и функционирование системы триотрофа – основной средообразующей консорции агробиоценоза на разных этапах органогенеза озимой пшеницы
Агробиоценозы представляют собой системы, образовавшиеся в результате сельскохозяйственной деятельности человека. Эти системы рассматриваются, как неустойчивые искусственные образования со слабо выраженной или разрушенной способностью к саморегуляции. Как и в структуре естественных биогеоценозов в них сформировались комплексы взаимосвязанных организмов, среди которых наибольшее значение имеют консорции, которые выделяют на основе трофических или топических отношений.
Представление о консорциях, как функциональной единице биогеоценозов, было сформулировано В.Н. Беклемишевым и Л.Г. Раменским, и развито в дальнейшем В.В. Мазингом. Консорцией называется совокупность видов, связанных пищевыми или прочими (топическими, фабрическими и др.) связями с некоторым видом, называемым эдификатором или детерминантом консорции, в качестве которого обычно выступает растение автотроф. Для любой консорции характерно то, что каждый вид занимает в сообществе особое место. Это обусловлено длительным приспособлением различных организмов к совместному проживанию в условиях достаточно динамичной среды. Каждый вид в сообществе занимает определенную экологическую нишу (совокупность условий обитания вида в сообществе и в системе взаимоотношений между видами и в использовании ресурсов).
Агробиоценоз озимой пшеницы представлены рядом консорций, детерминантом которых, в первую очередь, выступают растения самой культуры, а так же различные виды сорных растений.
В агробиоценозе озимой пшеницы достаточно четко просматриваются связи детерминанта (культура, иногда сорные растения) и разнообразных организмов – фитофагов, составляющих первый концентр (клоп-черепашка, злаковые тли, пшеничный трипс, хлебные пилильщики) и энтомофагов – второй концентр (хищники: кокцинеллиды, сирфиды, златоглазки, полосатый трипс и паразиты: теленомины, фазии, афидииды, коллирия), тесно связанные друг с другом в их жизнедеятельности (рис. 4). Это обусловливает поддержание определенного уровня потоков энергии и круговорот питательных веществ, не создавая условий для их концентрации в каких-то блоках, и повышая их целесообразную эффективность в пределах отдельных консорций, а затем и всего агробиоценоза.
Без выделения и изучения консортов в системе консорции озимой пшеницы наше представление о функционировании агробиоценоза этой культуры не будет полным. Агробиоценоз пшеницы представлен большим числом различных видов организмов, имеющих между собой самые разнообразные взаимоотношения. В структуре изучаемой нами консорции (триотрофа) основную роль играют трофические связи между ее составляющими, отражающими основное направление потока энергии от детерминанта — посева пшеницы и до пользователей последующих уровней – фитофагов озимой пшеницы, а от них – к зоофагам. В процессе изучения консортивных связей в посевах озимой пшеницы вырисовалась схема взаимодействия основных организмов первого и второго консортов (рис. 2).
В агроценозе озимой пшеницы между автотрофами: растениями озимой пшеницы (детерминантом) и различными видами сорных растений складываются конкурирующие отношения. Растения между собой конкурируют за основные источники жизни на первых этапах развития.

Рисунок 4 ‒ Основные консорты посева озимой пшеницы
Это почвенная влага и питательные вещества, в дальнейшем при нарастании вегетативной массы присоединяется еще один фактор – борьба за солнечную энергию – свет. Если консорция озимой пшеницы занимает детерминирующие положение, то консорции сорных растений в разных условиях увлажнения, плодородия почвы, её аэрации и др. абиотических факторов подвержены варьированию в пределах одного ландшафта и, как правило, характеризуются сходством формирования консортивных связей с основным детерминантом.
Автотрофы (озимая пшеница, занимающая доминирующие положение, иногда сорные растения) обеспечивают консорты первого концентра энергией, поэтому взаимоотношения этих организмов определяются их пищевыми интересами. Кроме того, фитофаги, осуществив механическое
нарушение каких-либо тканей у растений пшеницы, вводят в них свои выделения, что облегчают освоение растений бактериями и грибами.
Автотрофы выступают также и как средообразующий фактор для фитофагов. В зависимости от того, в какой стадии органогенеза находится пшеница, консорты первого порядка питаются различными её органами, как правило, теми, где содержится наибольшее количество пластических веществ.
Известно, что фитофаги характеризуются строгой приуроченностью питания на определенных органах растений. Так при питании на озимой пшенице одни фитофаги предпочитают питаться на колосе (антофаги – обитатели генеративных органов или капрофаги – повреждают зерновки), другие – листья (филлофаги), третьи – стебель (каулофаги). Есть фитофаги, использующие в качестве источника пищи сначала листья я, переходят затем и на колос.
Весной в фазу трубкования — цветения часть фитофагов (щитники-черепашки, злаковые тли) питаются на листьях открыто, в связи с чем являются открыто вредящими видами консортов первого концентра, что обусловило круг их паразитов и хищников, то есть консортов второго концентра.
Другие виды фитофагов развиваются во влагалище листа (пшеничный трипс), третьи — внутри стебля (хлебные пилильщики) и т. д. По мере формирования и созревания зерновок в колосе, основная масса фитофагов (щитники-черепашки, злаковые тли, пшеничный трипс) переходит к питанию на нем, а хлебный пилильщик опускается вниз внутри стебля пшеницы. В зависимости от образа жизни и распределения питательных веществ, происходит вертикальная миграция консортов первого концентра в агроценозе пшеничных посевов.
Все изученные нами консорты первого концентра детерминант обеспечивает энергией и веществами, а также является субстратом для их обитания или временного прикрепления. Гетеротрофы первого концентра используют энергию и вещества, находящиеся в живых тканях автотрофа — в стебле, листьях, соцветиях, зерне и являются биотрофами.
Биотрофы первого порядка потребляют органические вещества и обеспечивают их частичную минерализацию, то есть, являются отчасти редуцентами. Таким образом, биотрофы представляют собой достаточно обособленные территориально и функционально группы организмов.
Связанные с ними консорты второго концентра также образуют относительно обособленные группы. Зоофаги имеют более сложную структуру, разделяясь на биотрофов-хищников и биотрофов-паразитов, которые в свою очередь делятся на специализированных (монофагов), то есть приуроченных к одному виду консорта первого концентра, и неспециализированных – олигофагов и полифагов.
Зоофаги вредной черепашки относятся к биотрофам-паразитам, но делятся на специализированных (мухи-фазии), которые тесно связаны с циклом развития хозяина и его численностью, и неспециализированных теленомин, которые в течение откладки яиц вредной черепашкой развиваются в трех поколениях, а после перелетают в посевы других культур, где обитают бивальтинные клопы.
Зоофаги злаковых тлей делятся на биотрофов-хищников и биотрофов — паразитов. У тлей мы не наблюдали наличия специализированных консортов второго концентра. И хищники, и паразиты являются либо полифагами (концинеллиды, златоглазки), либо олиготрофами (сирфиды, афидииды). Кокцинеллиды и златоглазки на озимой пшенице нападают не только на злаковых тлей, но и на других мелких насекомых, но основной их пищей в агроценозе являются тли. Сирфиды и афидииды в консорции агроценоза пшеницы питаются только злаковой тлей, но при достижении детерминантом полной спелости переходят в соседние консорции культур, где питаются тоже тлями других видов.
Такие консорты первого концентра, как пшеничный трипс и обыкновенный хлебный пилильщик являются скрытноживущими видами, что и обусловило состав их зоофагов, которые являются специализированными фагами этих фитофагов. Полосатый трипс (Aelothrips fasciatus L.) развивается синхронно со своей жертвой — пшеничным трипсом, прилетая с ним в одно время на посевы культуры, питаясь им и также уходя на зимовку одновременно со своей жертвой.
Коллирия (Collyria coxator Vill) является узкоспециализированным зоофагом обыкновенного хлебного пилильщика, который заражает яйца фитофага и, проходя сопряжено все стадии развития личинки с консортом первого порядка в его теле, убивает его только весной следующего года.
Зоофаги выделяются избирательностью в образе жизни в зависимости от особенностей биологии консорта первого концентра: биотрофы скрытноживущих фитофагов являются специализированными паразитами и хищниками, зоофаги открыто живущих фитофагов в большей массе являются полифагами и олигофагами.
Формирование и становление основной трофической консории агробиоценоза озимой пшеницы – триотрофа в условиях неустойчивого увлажнения Ставрополья осуществлялось в течение длительного времени в результате сопряженного развития продуцента и консументов первого и второго порядков (в основном фитофагов и их зоофагов).
Поскольку все три составляющие триотрофа способны совместно существовать в определенных условиях экотипа, то естественно, что они являются экологически совместимыми. Становление консортов пшеницы шло в прошлом и поддерживается в настоящее время в основном за счет местной фауны (Глазунова, 2004а).
Состояние и активность компонентов триотрофа периодически изменяются, что оказывает определенное влияние на различные стороны взаимоотношения между ними.
Кормовое растение, являясь в системе триотрофа продуцентом, влияет на следующее его звено – консументов первого порядка – фитофагов, определяя, в зависимости от качества пищи, их жизнеспособность и устойчивость к различным внешним воздействиям, в том числе и к энтомофагам. В свою очередь, физиологическое состояние фитофагов влияет на степень их поражения консументами второго порядка – энтомофагами.
Таким образом, экологические связи в системе триотрофа определяют численность и популяционную динамику фитофагов с учетом влиянием на них с одной стороны кормового растения, с другой — энтомофагов. Рассмотрим эти взаимодействия на примере доминантных видов вредителей озимой пшеницы, концентрирующихся на ней и причиняющих вред в весенне-летний период ее онтогенеза (VI-XI этапы органогенеза).
4.2 Изучение и описание биологических особенности развития фитофагов, наносящих вред в период формирования репродуктивных органов озимой пшеницы
Вредная черепашка и её энтомофаги. Этот вредитель озимой пшеницы занимает особое положение среди выявленных массовых видов фитофагов, так как выделяется из этой группы как супердоминат, поскольку многие годы сохраняет свою численность выше ЭПВ, расширяет свой видовой ареал и ареал вредоносности, утратил многолетнюю ритмику развития.
Наши исследования, показали, что в сборах насекомых встречалось 8 видов клопов (рис.5). Четыре вида относились к сем. Scutelleridae (Щитники-черепашки) – Eurygaster integriceps Put., Eurygaster maura L., Eurygaster austriacus Schr., Erygaster testudinaris L., и четыре вида относились к сем. Pentatomidae (Щитники) – Aelia acumunata L., Aelia rostrata L., Carpocoris fuscispinus Boh., Dolycoris baccarum L. Все года в наших учетах преобладала вредная черепашка. Ее численность составила в среднем — 79,2 % от общего количества собранных клопов (рис. 5). По вредоносности этот вид также превосходит другие виды, так как снижает не только количество, но и качество урожая.
Долгосрочная взаимосвязь онтогенеза вредной черепашки и озимой пшеницы привела к четко выраженной приуроченности питания клопов в зоне локализации конуса нарастания и по мере роста и развития растения, места питания фитофага соответственно меняются. Так на II этапе органогенеза пшеницы клоп питается в зоне эмбрионального колоса, на V-VII этапах – в зоне формирующегося колоса, после выколашивания растений – на колосе.
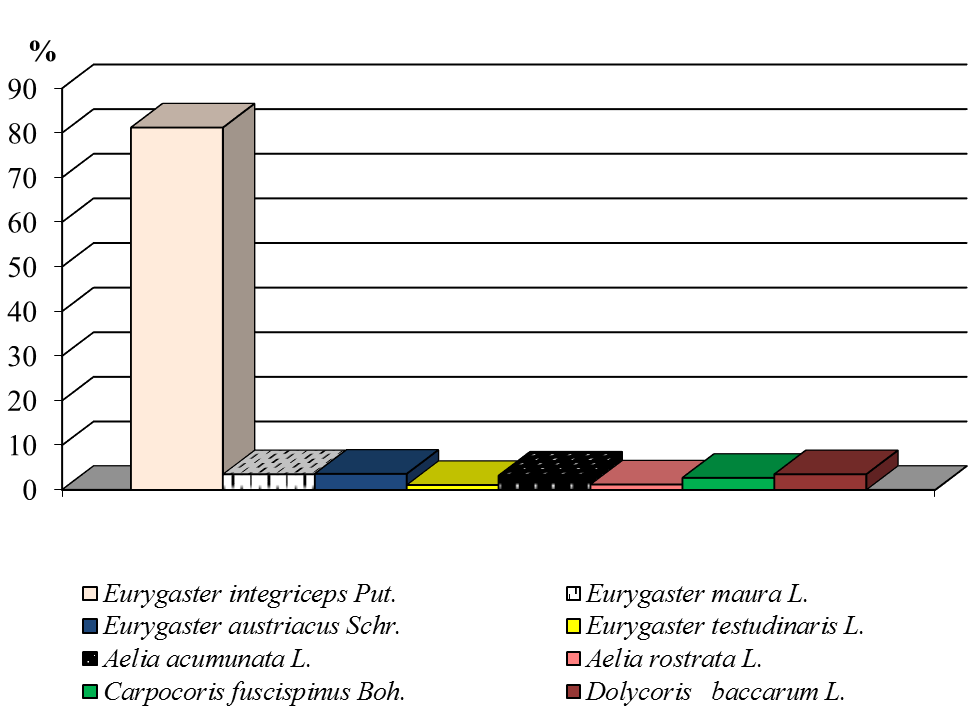
Рисунок 5 ‒ Виды клопов семейства Scutelleridae и Pentatomidae и их доля в посевах озимой пшеницы по годам исследования
Это объясняется тем, что клопы питаются в зоне, где идет усиленный приток транспортных форм групп биополимеров, который лучше усваивается в их организме.
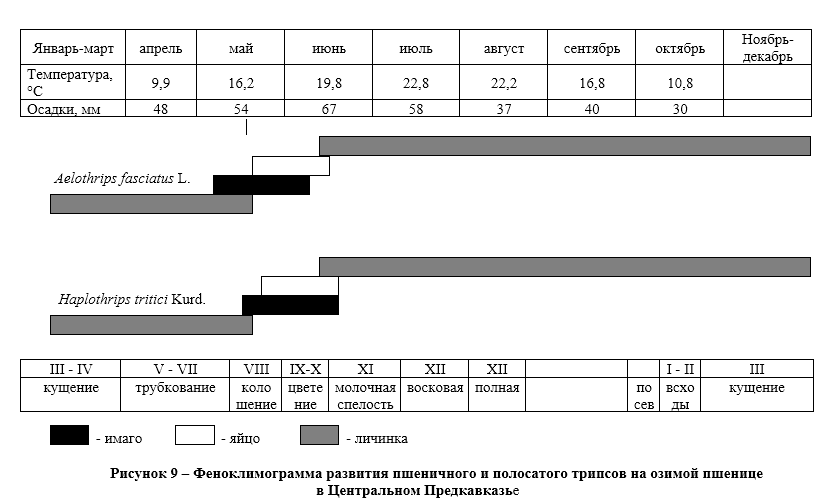
В Центральном Предкавказье вредная черепашка появляется на полях озимой пшеницы в начале третьей декады апреля, когда растения озимой пшеницы находятся на VI-VII этапах органогенеза, т.е. в фазе трубкования (рис. 6). Перезимовавшие клопы встречались нами в посевах до конца июня,
Наблюдения показали (рис. 6), что начало откладки яиц вредной черепашкой отмечено в первой декаде мая, а массовая откладка яиц с начала второй декады мая – по вторую декаду июня. Завершается она к концу июня, что соответствует VII-ХI этапам органогенеза.
.
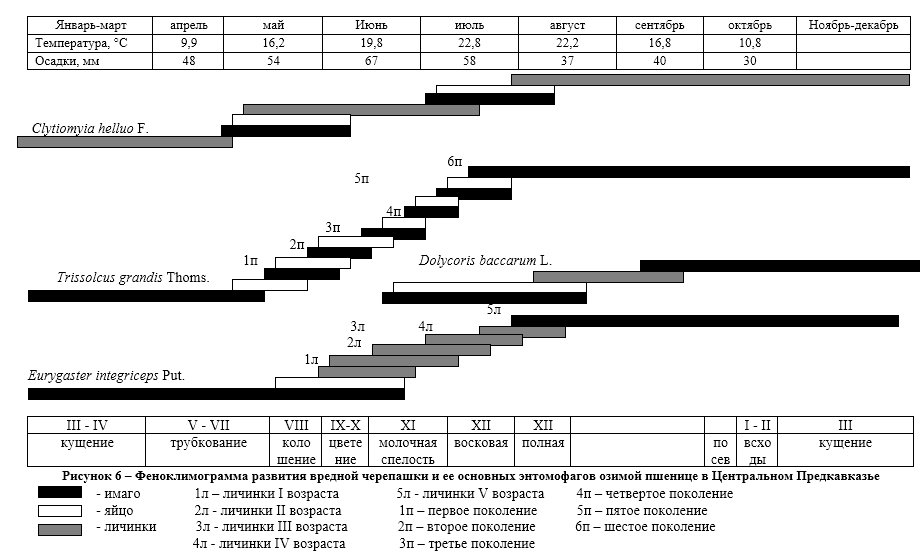
Личинки первого возраста начинают появляться в конце мая – начале июня, массовое отрождение личинок наблюдается с первой и вторую декады июня, растения в этот период находятся на IХ – ХI этапах органогенеза. Личинки второго возраста в массе наблюдались во второй и третьей декаде июня, когда растения озимой пшеницы достигают на Х-ХI этапа органогенеза. Личинки третьего и четвертого возрастов в посевах наблюдались с третьей декады июня, когда растения озимой пшеницы находятся на ХI этапе органогенеза (фаза молочной спелости зерна).
Личинки пятого возраста и молодые имаго в массе начинают появляться в первой декаде и до начала второй декады июля, при переходе растений на ХII этап органогенеза (фазы восковой и полной спелости зерна). Перелет в места зимовки наблюдается после уборки озимой пшеницы в конце второй – начале третьей декады июля (рис. 6).
Перезимовавших клопы на посевах озимой пшеницы питается в зоне формирующегося колоса (VI-VII этапы органогенеза), который в связи с интенсивным ростом быстро перемещается кверху. Наносимые клопами повреждения растениям в этот период приводит к деформации или остановки развития формирующегося колоса, вызывая частичную белоколосость
При наступлении VIII этапа органогенеза (фаза колошения) появляется колос и имаго клопов переходят к питанию на нем, вызывая белоколосость и изгиб колосоножки. Такие же симптомы и вредоносность фитофага мы отмечали и на IХ этапе органогенеза (фаза начала цветения). При переходе растений на Х — ХI этап органогенеза (цветение-начало роста и формирования зерновки) питание клопов вызывает череззерницу и недоразвитость, щуплость зерновок.
Таким образом, питание перезимовавших имаго вредной черепашки на растениях озимой пшеницы в Центральном Предкавказье приводит к количественным потерям урожая культуры.
Известно, и нашими исследованиями подтверждается, что личинки вредной черепашки первого возраста не питаются. Питание личинок остальных возрастов отмечено на формирующемся зерне колоса. Вредоносность и симптомы повреждения зависели от этапа органогенеза озимой пшеницы.
При питании личинок вредной черепашки на Х этапе органогенеза отмечали череззерницу; на ХI этапе – недоразвитость, (щуплость) зерновок; на ХII этапе – щуплость зерновок и снижение массы 1000 зерен.
Итак, питание личинок и имаго молодых клопов на растениях приводит к количественным потерям урожая культуры (недоразвитость, щуплость зерновок и снижение массы 1000 зерен), а так же к качественным потерям, так как при повреждении 2-3 % зерен вредной черепашкой, наблюдается существенное снижение технологических и хлебопекарных качеств культуры.
Численность вредной черепашки ограничивают яйцепаразиты – теленомины. Мы собирали зараженные этим паразитами кладки яиц вредной черепашки и в лабораторных условиях определяли степень заражения и видовой их состав.
Среди выведенных паразитов все годы исследования доминировал Trissolcus grandis Thorns. В среднем за годы исследований этот паразит заражал до 48,2 % яиц клопа-черепашки. Второе место по эффективности занимал Telenomus chloropus Thorns. (29,3 %), третье место — Ooencirtus telenomicida Vass. (14,5 %). Остальные виды паразитов встречались в незначительных количествах (рис. 7).
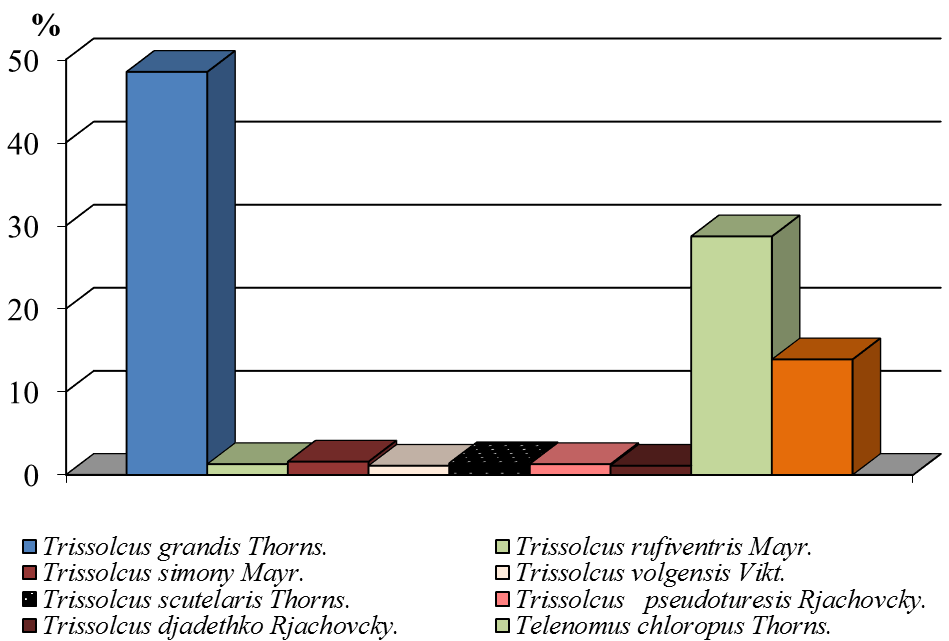
Рисунок 7 ‒ Доля видов сем. Scelionidae и Encyrtidae в посевах озимой пшеницы
В природно-климатических условиях Центрального Предкавказья в яйцах вредной черепашки на посевах озимой пшеницы развивается три поколения яйцеедов. Вылет перезимовавших теленомин начинается в первой половине апреля. Сначала они концентрируются на опушках лесополос, где питаются нектаром цветущих дикорастущих трав. Позднее, начиная с средины второй декады апреля, перелетают на посевы озимой пшеницы, когда не посевах появляются клопы вредной черепашки и начинают откладывать яйца. Первое поколение теленомин развивается в среднем 24-28 дней, вылет имаго происходит в конце третьей декады мая. Второе поколение развивается 16-18 дней, вылет имаго приходится на середину второй декады июня, а третье поколение развивается за 12-14 дней и имаго вылетает в третьей декаде июня (рис. 6).
Возможность дальнейшего размножения теленомин после завершения развития третьего поколения на вредной черепашке, определяется присутствием на пшенице и в других стациях дополнительных хозяев. В начале июля теленомины мигрируют на пропашные культуры и в природные стации, где заражают яйца второго поколения бивольтинных клопов.
Значительную роль в ограничении численности вредной черепашки играют мухи-фазии (Diptera, Tachinidae, Phasiinae). Они являются вторыми по значению, после теленомин, паразитами вредителя.
Паразиты имаго вредной черепашки в нашем регионе представлены тремя видами мух-фазий: золотистой (Clytiomyia helluo F.), серой (Alophora subcoleoptrata L.) и пестрой (Phasia (Ectophasia) crassipennis F.). На посевах озимой пшеницы более многочисленна и постоянно доминирует золотистая фазия (51 %), несколько реже встречается серая (44 %), пестрая фазия была малочисленна (5%) (рис. 8).
В климатических условиях Центрального Предкавказья золотистая и пестрая фазии развиваются на черепашке в двух поколениях. По нашим наблюдениям они характеризуются значительным сходством сезонных циклов развития. С 2001года мы наблюдали за фенологией этих паразитов.
У золотистой и пестрой фазий зимует личинка второго возраста в молодых клопах. В конце апреля — начале мая личинки выходят из тела клопов и окукливаются в почве. Первые мухи появляются в начале первой декады мая и после непродолжительного дополнительного питания (3-4 дня) приступают к откладке яиц на вредную черепашку. Золотистая фазия первого поколения откладывает яйца на глаза клопов. Период яйцекладки продолжается до конца июня, что совпадает с откладкой яиц вредной черепашки. Личинки фазий первого поколения покидают клопов для окукливания в почве с начала третьей декады июня по середину второй декады июля. К этому времени в агроценозе озимой пшеницы в массе появляются личинки IV-V возрастов вредной черепашки. Взрослые мухи второго поколения начинают вылетать с середины третьей декады июня — до конца первой декады июля перед началом окрыления молодых клопов и заражают их до отлета в места зимовки. Все развитие фазий первого поколения в условиях зоны неустойчивого увлажнения длится 30-40 дней, в зависимости от погодных условий года.
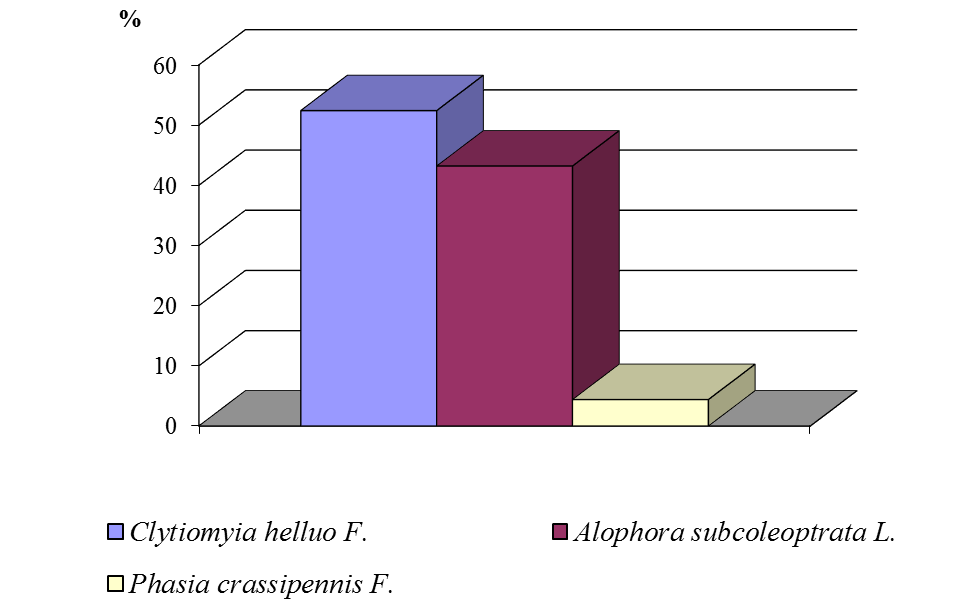
Рисунок 8 ‒ Доля видов сем. Tachinidae в посевах озимой пшеницы
В отличие от золотистой и пестрой фазий, серая фазия зимует вне хозяина, в виде пупария в почве. Этот вид фазий более тесно связан с природными стациями. На посевных озимой пшеницы развивается только летнее поколение паразита. Развитие второй генерации происходит в местах зимовки вредной черепашки – лесополосах.
Взрослые мухи вылетают рано весной, обычно в середине или конце второй декады апреля и заражают клопов еще в местах зимовки. С перелетом клопов на поля серая фазия мигрирует за черепашкой и продолжает ее заражать до середины мая. Из мест зимовки зараженные клопы, как и здоровые, перелетают на озимые, но практически не откладывают яйца из-за стерилизации самок.
Развитие серой фазии от личинки I возраста до окукливания в разные годы длится от 40 до 55 дней. Куколки появляются во второй декаде июля. Первые мухи серой фазии появляются в конце второй декады июля и присутствуют в посевах озимой пшеницы до уборки, заражая молодых клопов и личинок IV-V возраста. Возможность заражения клопов на полях ограничена во времени, но из-за неодновременного окрыления и отлета вредителя, число паразитированных серой фазией особей достигает 13%. Зараженные клопы, как и здоровые, перелетают в места зимовки. Личинки мух к концу августа достигают III возраста и выходят из хозяина в почву, где образуют пупарий. Хозяин вскоре погибает.
Таким образом, фазии снижают численность вредной черепашки, уничтожая молодых клопов осенью и приводя к стерилизации самок весной.
Пшеничный трипс и его энтомофаги. В последние десятилетия в Центральном Предкавказье, в частности в Ставропольском крае, отмечено нарастание численности и вредоносности пшеничного трипса.
Зимует у пшеничного трипса личинка старшего возраста — пронимфа в почве на полях озимой пшеницы прошлого года (рис.9).
Весной, когда наступает сухая и теплая погода в конце третьей декады апреля — начале первой декады мая, личинки выходят на поверхность и превращаются в нимф. Имаго пшеничного трипса появляются в первой декаде мая, кормовое растение в этот период находится на VII этапе органогенеза (фаза выхода в трубку). Интенсивный лет трипсов приходиться на третью декаду мая и совпадает с массовым колошением озимых – VIII этапом органогенеза.
Отродившиеся трипсы приступают к питанию соком на растущих молодых тканях растения, чаще всего это формирующийся колос или формирующийся флаг лист. При питании трипсов на VIII этапе органогенеза озимой пшеницы наблюдается деформация (скручивание и гофрирование) флагового листа и остей колоса, обесцвечивание листьев в местах питания насекомых. На IХ-Х этапах органогенеза озимой пшеницы трипсы переходят питаться на колос, в результате чего поврежденные колосья могут деформироваться, их вершина становится рыхлой, и на вид растрепанной, может отмечаться частичная белоколосость и пустоцветность. Если предшественником была озимая пшеница, то пшеничный трипс наносит вред посевам раньше – на VI этапе органогенеза в период выхода растений в трубку, и еще до начала колошения проникает в колос через флаг-лист. При большой численности они причиняют значительный вред еще не выколосившемуся растению.
После перелета на посевы озимой пшеницы через неделю приступают к спариванию и откладке яиц. Откладка яиц у трипсов в климатических условиях Центрального Предкавказья по нашим исследованиям длится в среднем около месяца – с третьей декады мая по третью декаду июня. Эмбриональное развитие длится около недели в оптимальных условиях, при пониженных температурах до десяти дней. Массовое отрождение личинок приходится на конец второй начала третьей декады июня, когда растения озимой пшеницы достигают ХI этапа органогенеза (фаза молочной спелости зерна). Личинки в среднем развиваются около двадцати дней.
Наиболее вредоносны личинки, которые питаются на молодых зерновках озимой пшеницы (ХI-ХII этапы органогенеза). В местах питания трипсов на зерновках появляются мелкие желто-бурые пятна, зерно становится щуплым, может деформироваться, снижается масса 1000 семян. К моменту уборки (конец июля) основная масса личинок заканчивает питание, и уходит в почву, подготавливаясь к зимовке.
Хлебные пилильщики. Опасными вредителями озимой пшеницы для зоны Центрального Предкавказья, а особенно в Ставропольском крае являются стеблевые хлебные пилильщики. По данным Е.В. Ченикаловой и В.Е. Чернова в различных природно-климатических зонах Ставропольского края они снижают урожай на 2,5-4 центнеров с одного гектара озимой пшеницы, что наносить ощутимый вред валовому сбору урожая в регионе.
Внутри соломины личинки пилильщиков съедает паренхиму и заполняет её экскрементами. На ХII этапе органогенеза озимой пшеницы пилильщики в нижнем междоузлие на уровне почвы делают кольцеобразный надрез, в результате чего основная масса стеблей надламывается и полегает до уборки урожая.
В условиях зоны Центрального Предкавказья в посевах озимой пшеницы встречается два вида стеблевых хлебных пилильщиков – обыкновенный хлебный пилильщик (Cephus pygmaeus L.) и черный хлебный пилильщик (Trachelus tabidus F.). Преобладающим видом в годы наблюдений был обыкновенный хлебный пилильщик, доля которого в наших сборах составляла 54,6-58,1 %, что в среднем на 15% больше, чем доля черного пилильщика.
Обыкновенный хлебный пилильщик в посевах озимой пшеницы появляется в конце первой начале второй декады мая в зависимости от погодных условий. Обычно его лет совпадает с началом колошения озимой пшеницы (VII этап органогенеза) (рисунок 10).
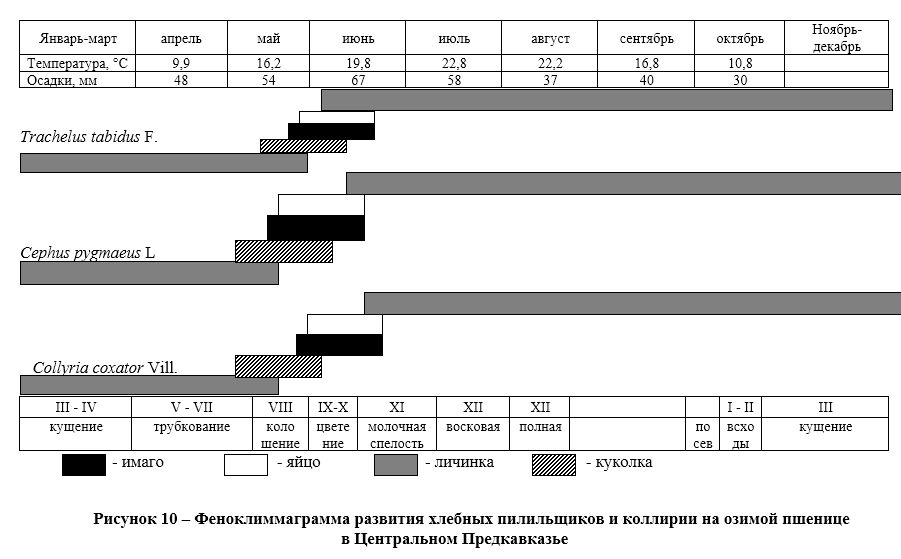
Массовый лет у обыкновенного хлебного пилильщика нами отмечен с середины второй декады мая до начала второй декады июня, что соответствует VIII-ХI этапу органогенеза озимой пшеницы.
Черный хлебный пилильщик вылетает на декаду позже обыкновенного в конце третьей декады мая в начале первой декады июня, когда озимая пшеница находиться на IХ этапе органогенеза озимой пшеницы или фазе цветения. Массовый его лет проходит в Центральном Предкавказье с начала первой декады и продолжается до конца второй декады июня, что соответствует IХ-ХI этапам органогенеза озимой пшеницы. После непродолжительного питания от трех до
пяти дней на нектароносной растительности, происходит спаривание пилильщиков.
Обыкновенный хлебный пилильщик откладывает яйца с VIII по ХI этап органогенеза, черный – с IХ по ХI этап органогенеза озимой пшеницы в верхнее междоузлие. Личинки начинают отрождаться в первой декаде июня последние личинки первою возраста встречались нами до конца второй декады июня. Личинки второго возраста начинают появляться с начала второй декады июня и встречаются до начала второй декады июля. Переход личинок в третий возраст был отмечен с начала третьей декады июня. В конце июня личинки пилильщиков начинают продвигаться по соломине к нижнему междоузлию. Этот процесс происходит постепенно, и к уборке их основная масса достигает первого междоузлия, которое и подпиливает. Развитие личинок приходиться на ХI- ХII этапы органогенеза озимой пшеницы. Питание личинок приводит к недоразвитию колосков и образованию щуплого неполновесного зерна. В растениях, где живет и развивается личинка стеблевого хлебного пилильщика колос и верхнее междоузлие имеют белесый цвет.
Злаковые тли и её афидофаги. Злаковые тли постоянно присутствуют в агробиоценозе озимой пшеницы. По нашим исследованиям, в зоне неустойчивого увлажнения Ставропольского края наибольшее хозяйственное значение среди злаковых тлей имеют большая злаковая тля (Sitobion avenae F.) и обыкновенная злаковая тля (Schizaphis graminum Rond.). Сопряженное развитие консументов с продуцентом в системе триотрофа представлено на рисунках 11-12.
Самки-основательницы отрождаются на посевах озимой пшеницы в третьей декаде апреля – начале первой декаде мая, когда озимая пшеница находиться на VII-VIII этапах органогенеза. Самки основательницы располагаются на флаговом листе образуя колонии. В результате питания злаковых тлей наблюдалось обесцвечивание листовой пластины при разрастании колонии отмечали скручивание и отмирание флагового листа. В дальнейшем на Х-ХI этапах органогенеза озимой пшеницы злаковые тли переходят питаться на её колос. Питание на колосе приводит к формированию щуплого зерна и к уменьшению количества зерен в колосе. В мае развитие одной генерации тлей длится от 17 до 20 дней, летом в июне-июле от 8 до 13 дней.
В зоне наших исследований злаковые тли развиваются в 8 поколениях в агроценозе озимой пшеницы. После восковой спелости зерна данные вредители покидают агробиоценоз озимой пшеницы и перелетают на дикие злаки, где дают еще 3 поколения. В конце третьей декады сентября – до середины второй декады октября, когда озимая пшеница находиться на II-III этапах органогенеза, на ней развивается последнее – двенадцатое поколение тлей. На растениях пшеницы появляются самки – полоноски, рождающие личинок, превращающихся в крылатых самцов и бескрылых самок, которые откладывают зимующие яйца.
Видовой состав афидофагов злаковых тлей в зоне исследования насчитывает 36 видов насекомых, в том числе 29 хищников: жесткокрылые (Coleoptera, сем. Coccinellidae), двукрылых (Diptera сем. Syrphidae) и
сетчатокрылых (Neuroptera, сем. Chrysopidae) и 7 паразитов из отрядов перепончатокрылых (Hymenoptera, сем. Aphidiidae).
Афидииды. Из мумифицированных тлей нами было выведено 7 видов перепончатокрылых паразитов. Доминирующим видом среди них был Aphidius ervi Hal., на втором месте по зараженности тлей Aphidius picipes Nees. Третье место по степени заражения злаковых тлей занимал Praon volucre Hal., остальные виды встречались в единичных количествах (рис. 13).
По своей фенологии указанные паразиты близки. Афидииды, как правило, зимуют в фазе личинки последнего возраста или предкуколки в мумифицированной тле на посевах озимой пшеницы, озимого ячменя и на злаках в окружающих лесополосах.
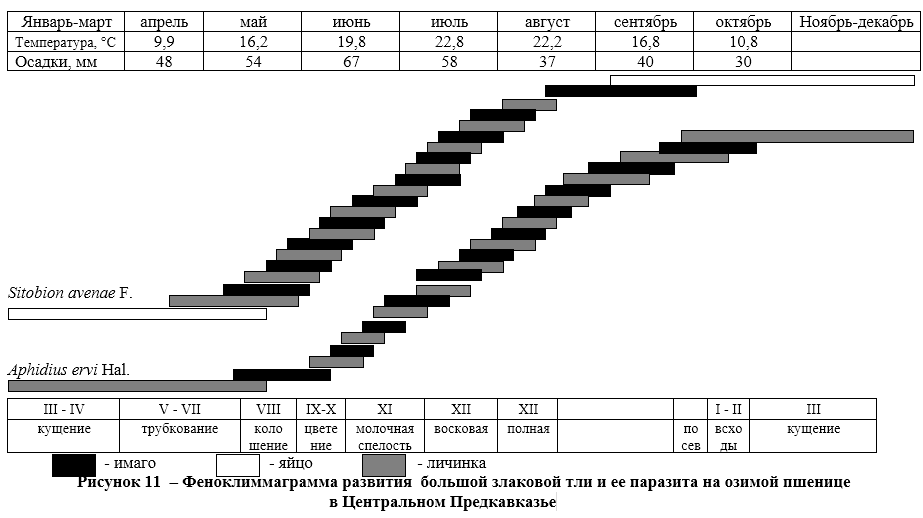
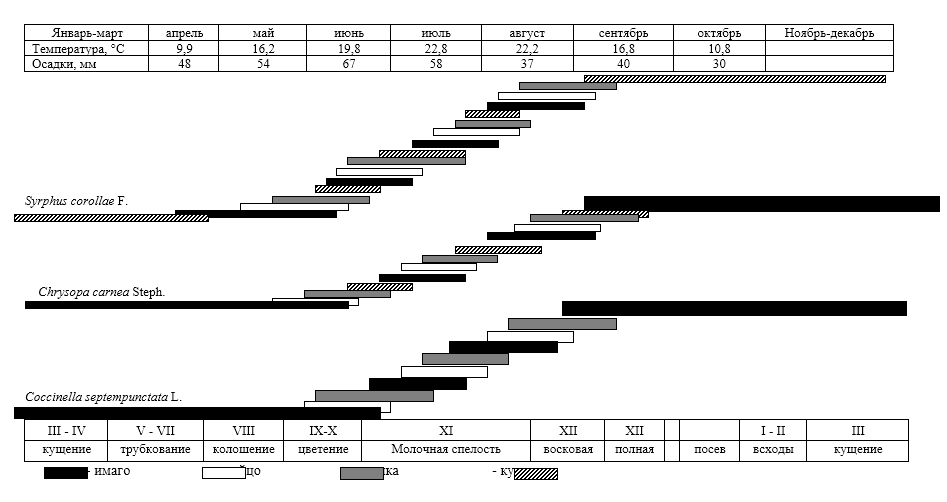
Рисунок 12 ‒ Феноклимаграмма развития хищников Syrphus corollae F., Chrysopa carnea Steph. и Coccinella septempunctata L. на озимой пшенице в Центральном Предкавказье
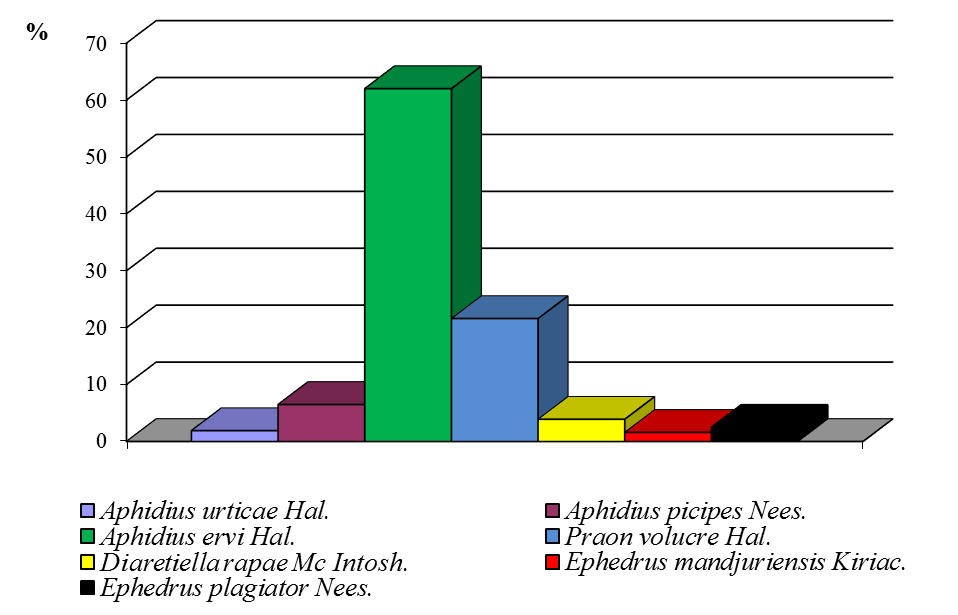
Рисунок 13 ‒ Доля видов сем. Aphidiidae в посевах озимой пшеницы
Весной в первой декаде мая появляются имаго афидиид, которые после спаривания откладывают яйца в имаго или личинок старших возрастов тли. Как показали наблюдения, первое поколение афидиид первое поколение афидиид развивается весной в течение 18-22 дней, в летние месяцы развитие сокращается до 12-14 дней, однако во время ливневых дождей и понижения температуры их развитие удлиняется. На популяции злаковых тлей в весенне-летний период в посевах озимой пшеницы развивается до 6 поколений афидиид. После ухода злаковых тлей
на зимовку они мигрируют на кукурузу и подсолнечник или дикорастущие растения, заселенные тлями. Осенью после появления всходов озимой пшеницы и перелета на них самок-полоносок тлей, туда же перелетает и часть популяции афидиид, которые заражают их и остаются зимовать в фазе личинки в теле тлей на посевах озимой пшеницы (рисунок 13).
Тлевые коровки. B агробиоценозе озимой пшеницы мы выявили 8 видов тлевых коровок, среди которых наиболее многочисленными были Coccinella septempunctata L. и Propylaea quatuordecimpunctata L. Эти виды составляли в среднем 86,1 % от общего числа собранных кокцинеллид (рис. 14). В зоне неустойчивого увлажнения кокцинеллиды развиваются в 3-х поколениях (рисунок 14), однако с теплой и сухой осенью, мы наблюдали развитие четвертого факультативного поколения.
Рисунок 14 ‒ Доля видов сем. Coccinellidae в посевах озимой пшеницы
В посевах озимой пшеницы до уборки успевает развиться 2 полных поколения кокцинеллид. Зимуют взрослые жуки, в основном, в подстилке лесополос, размещенных преимущественно в западном направлении. Поля озимой пшеницы кокцинеллиды начинают заселять в середине апреля, когда на них еще нет тлей. Имаго в этот период питаются нектаром цветущих растений. Самки начинают откладывать яйца в середине второй декады мая, когда на посевах появляются тли. Откладка яиц длится около месяца. Массовое отрождение личинок происходит в конце мая – начале июня. Личинки в своем развитии проходят в течение 17-21 дней 4 возраста. Закончив питание, личинки окукливаются открыто на растениях. Через 5-7 дней из куколок появляются взрослые жуки. Массовое отрождение жуков первого поколения наблюдается в конце июня.
Молодые жуки интенсивно питаются тлей и сразу же приступают к откладке яиц. В первой декаде июля начинается массовое отрождение личинок 2-й генерации. В конце июля – начале августа отрождаются жуки второго поколения. С посевов озимой пшеницы тлевые коровки мигрируют на посевы других культур заселенных тлями, где продолжают питаться и развиваться. Уходят на зимовку в середине октября.
Мухи-сирфиды (журчалки). Мухи этого семейства являются эффективными хищниками злаковых тлей. В агробиоценозе озимой пшеницы нами было выявлено 8 видов сирфид. Среди этих видов хищников первостепенная роль принадлежит трем видам: Syrphus corollae F., S. balteatus Deg. u Sphaerophoria scripta L., численность которых в среднем за годы исследований составляет 81 % среди выявленных видов (рисунок 15).
Сирфиды в Центральном Предкавказье развиваются в 4 генерациях, зимуют в фазе куколки в почве или под растительными остатками. Весной мухи вылетают из мест зимовки в середине апреля. Самки появляются неполовозрелыми и только после дополнительного питания нектаром и пыльцой цветущей растительности вокруг полей или в лесополосах, примерно через две недели приступают к откладке яиц (рис. 15).
На посевах озимой пшеницы сирфиды появляются во время образования на листьях колоний тлей – в третьей декаде мая.
Наиболее многочисленные виды Syrphus corollae F. и Syrphus balteatus Deg. развиваются в трех генерациях в посевах озимой пшеницы, развитие четвертой генерации происходит на подсолнечнике, кукурузе и других культурах, заселенных тлями. Период развития от яйца до имаго у сирфид сравнительно короткий 24-28 дней.
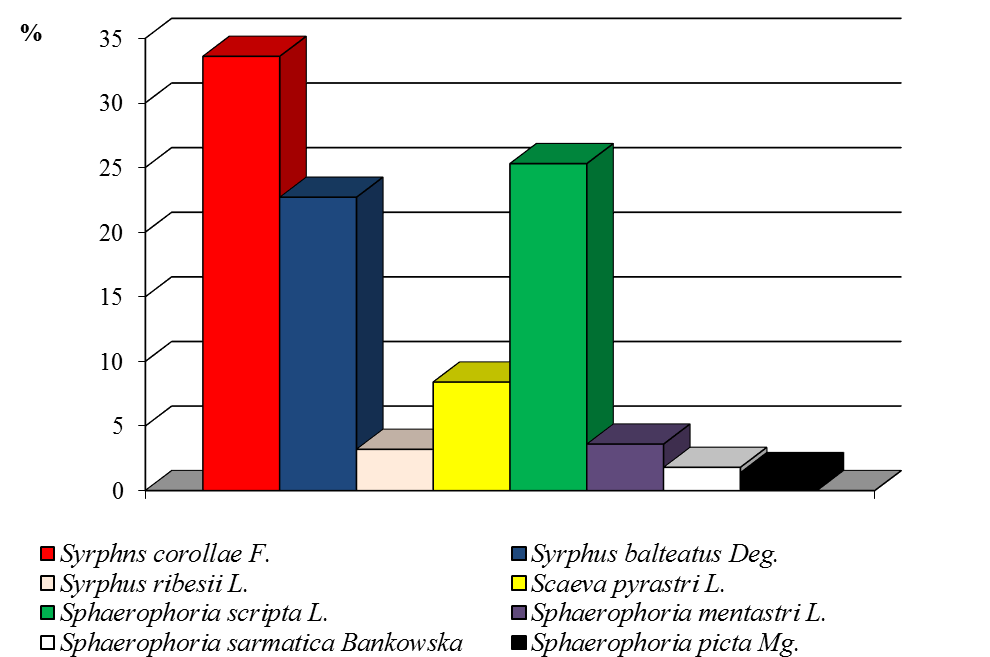
Рисунок 15 ‒ Доля видов сем.Syrphidae в посевах озимой пшеницы
по годам исследований
Наибольшее количество личинок первого поколения наблюдается с конца мая по середину июня. Мухи второго поколения появляются в посевах во второй половине июня. Массовое отрождение личинок второй генерации начинается с третьей декады июня и продолжается до конца первой декады июля. Имаго третьего поколения присутствуют на полях озимой пшеницы почти весь июль, личинки третьего поколения в основной массе заканчивают развитие к концу июля – началу августа. Четвертое поколение сирфид развивается вне агробиоценоза озимой пшеницы.
Вид Sphaerophoria scripta L. развивается в четырех поколениях, ее фенология сходна с фенологией Syrphus corollae F. и Syrphus balteatus Deg. Остальные виды рода Sphaerophoria развиваются аналогично, но предпочитают другие виды тлей. Одни виды в массе встречаются на овощных и плодовых культурах, другие виды – Syrphus ribesii L. и Scaeva pyrastri L., в нашей зоне развиваются только в двух поколениях, и лишь одно из них – в посевах озимой пшеницы. На зимовку сирфиды уходят в конце августа — начале сентября.
Златоглазки. 3a восемь лет исследований в агроценозе пшеничного поля нами было выявлено 3 вида златоглазок: обыкновенная (Chrysopa carnea Steph.), прозрачная (Chrysopa perla L,.) и красивая (Chrysopa formosa Br.). По обилию ведущее положение занимает златоглазка обыкновенная. В разные годы наблюдений она составила 78,9-92,3 % от общего числа выявленных видов златоглазок.
Имаго первого поколения златоглазок появляется в третьей декаде мая. Самкам златоглазки, как и самкам кокцинеллид, для созревания яиц необходима белковая пища. После не продолжительного питания тлями они начинают откладывать яйца. Эмбриональное развитие первого поколения длится около недели. Личинка за 11-15 дней проходит три возраста и окук-ливается. Развитие куколки продолжается до 18 дней. Массовое отрождение личинок первого поколения мы наблюдали в конце июня – начале июля в фазу молочно-восковой спелости озимой пшеницы. Имаго второго поколения появляются в конце июля и перелетают на другие культуры, заселенные тлями. Всего златоглазки развиваются в трех поколениях (рис. 12).
В результате проведенных наблюдений мы установили, что в массе доминантные виды фитофагов, причиняющие основной вред посевам озимой пшеницы в агробиоценозах Центрального Предкавказья, появляются на ней, начиная с VIII этапа органогенеза (фаза колошения). С этого этапа и по ХI этап органогенеза включительно (фаза молочной спелости) фитофаги питаются, спариваются и откладывают яйца, а так же появляются и развиваются их новые поколения (рисунок 16).
В условиях Центрального Предкавказья для развития озимой пшеницы с начала VIII этапа органогенеза (фаза колошения) до начала ХII этапа органогенеза (фаза восковой спелости зерна) в разные годы требуется от 28 до 35 дней. Именно в этот период вредители наносят ощутимый экономический вред посевам озимой пшеницы и культура нуждается в защите от их комплекса. На ХII этапе органогенеза растения озимой пшеницы грубеют и делаются не пригодными для питания вредителей. Это следует из анализа среднемноголетней динамики численности основных вредителей в процессе онтогенеза озимой пшеницы.
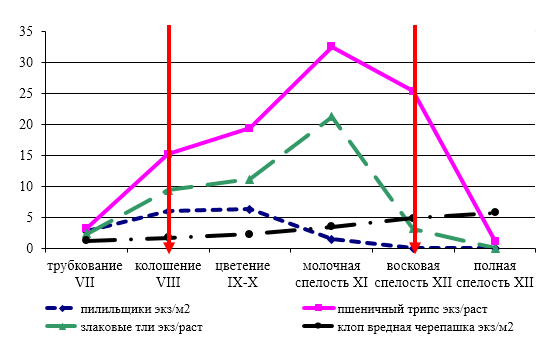
Рисунок 16 ‒ Изменение численности основных вредителей озимой пшеницы на разных этапах ее онтогенеза
(Ставропольский край, Шпаковский район)
Основу защитных мероприятий озимой пшеницы составляют химические препараты, применение которых сопровождает всю вегетацию культуры. Однако нарастает понимание значимости биологизации защиты растений, использования в первую очередь безопасных методов и средств борьбы с вредными организмами, производства продукции сельского хозяйства, не загрязненной токсикантами различного происхождения.
4.3. Определение биологической эффективности биоинсектицидов для различных групп фитофагов озимой пшеницы
В первый срок внесения инсектицидов предварительный учет численности насекомых проводили в фазу начала колошения озимой пшеницы перед ее обработкой препаратами. В эту фазу численность имаго вредной черепашки составляло 0,5 экз./м2 (ЭПВ перезимовавших клопов 1,5-3 экз./м2); злаковых тлей – 3 экз./растение (ЭПВ 5-10 экз./ растение при заселении выше 50% растений в фазы выхода в трубку – колошение); пшеничного трипса – 10 экз./растение (ЭПВ 40-50 экз./ растение в фазы от колошения до налива зерна); хлебных пилильщиков – 5 экз. на 100 взмахов сачка (ЭПВ по имаго 7-10 экз. на 100 взмахов сачка); пьявицы красногрудой – 1,2 экз./растение (ЭПВ 1 экз./ растение). Как видим по результатам учетов на момент первой обработки экономический порог вредоносности по численности превышала только пьявица красногрудая (таблица 5).
После обработки биоинсектицидами учет численности насекомых проводили через 3 дней 24 мая 2019 года. Озимая пшеница находились в этот момент в фазе колошения. В контроле численность вредной черепашки и пшеничного трипса осталась на прежнем уровне, увеличилась численность злаковых тлей до 4,2 экз./растение, хлебных пилильщиков до 6,4 на экз./ 10 взмахов сачка, пьявицы красногрудой до 1,4 экз./растение. В варианте опыта с применением биоинсектицида Биослип БВ с нормой расхода 3 л/га, наибольшая биологическая эффективность нами отмечена в отношении злаковых тлей и вредной черепашки – она составила 64 и 58 % соответственно, наименьшая эффективность отмечена в отношении хлебных пилильщиков их численность снизилась всего на 26 %.
В варианте опыта с применением биоинсектицида Биослип БТ с нормой расхода 3 л/га, наибольшая биологическая эффективность нами отмечена в отношении пьявицы красногрудой – она составила 75 %, наименьшая эффективность отмечена в отношении злаковых тлей и хлебных пилильщиков – 30 и 25 % соответственно. В варианте опыта с применением баковой смеси биоинсектицидов Биослип БВ и Биослип БТ с нормами расхода по 1,5 л/га, биологическая эффективность против вредной черепашки, пшеничного трипса злаковых тлей и пьявицы красногрудой колебалась в пределах 45-55 %, наименьшая эффективность отмечена в отношении хлебных пилильщиков их численность снизилась всего на 25 %.
Таблица 5 — Изменение численности вредителей в агробиоценозе озимой пшеницы при применении биоинсектицидов в фазу колошения
(учебно-опытная станция, сорт Юка, 2019 г.)
| Объекты
наблюдения |
Число до обработки | Количество по суткам учетов после обработки | Биологическая эффективность, % | |||
| 3 | 7 | 3 | 7 | |||
| Биослип БВ (3 л/га) | ||||||
| Вредная черепашка*** | 0,5 ± 0,05 | 0,21± 0,01 | 0,15± 0,01 | 58 | 70 | |
| Злаковые тли* | 3 ± 0,2 | 1,1± 0,1 | 0,8± 0,1 | 64 | 75 | |
| Пшеничный трипс* | 10 ± 0,8 | 5,5± 0,4 | 4,2± 0,3 | 45 | 58 | |
| Хлебные ** пилильщики | 5 ± 0,5 | 3,7± 0,3 | 3,4± 0,3 | 26 | 32 | |
| Пьявица красногрудая* | 1,2 ± 0,1 | 0,7± 0,01 | 0,8± 0,01 | 42 | 37 | |
| Биослип БТ (3 л/га) | ||||||
| Вредная черепашка*** | 0,5 ± 0,05 | 0,29± 0,01 | 0,25± 0,01 | 43 | 51 | |
| Злаковые тли* | 3 ± 0,2 | 2,1± 0,1 | 2,0± 0,1 | 30 | 35 | |
| Пшеничный трипс* | 10 ± 0,8 | 5,4± 0,4 | 6,0± 0,4 | 46 | 40 | |
| Хлебные ** пилильщики | 5 ± 0,5 | 3,8± 0,3 | 3,5± 0,3 | 25 | 30 | |
| Пьявица красногрудая* | 1,2 ± 0,1 | 0,3± 0,01 | 0,2± 0,01 | 75 | 85 | |
| Биослип БВ + Биослип БТ (1,5 + 1,5 л/га) | ||||||
| Вредная черепашка*** | 0,5 ± 0,05 | 0,24± 0,01 | 0,23± 0,01 | 52 | 55 | |
| Злаковые тли* | 3 ± 0,2 | 1,7± 0,1 | 1,6± 0,1 | 45 | 48 | |
| Пшеничный трипс* | 10 ± 0,8 | 4,5± 0,4 | 4,0± 0,3 | 55 | 60 | |
| Хлебные ** пилильщики | 5 ± 0,5 | 3,8± 0,3 | 3,5± 0,3 | 25 | 30 | |
| Пьявица красногрудая* | 1,2 ± 0,1 | 0,6± 0,01 | 0,5± 0,01 | 50 | 55 | |
| Контроль | ||||||
| Вредная черепашка*** | 0,5 ± 0,05 | 0,5 ±0,05 | 0,7±0,05 | — | ||
| Злаковые тли* | 3 ± 0,2 | 4,2± 0,4 | 5,6± 0,5 | — | ||
| Пшеничный трипс* | 10 ± 0,8 | 10 ± 0,8 | 10 ± 0,8 | — | ||
| Хлебные ** пилильщики | 5 ± 0,5 | 6,4±0,5 | 7,2±0,5 | — | ||
| Пьявица красногрудая* | 1,2 ± 0,1 | 1,4±0,1 | 1,6± 0,2 | — | ||
Примечание:*-численность на экз./колос (растение) **- численность на экз./ 10 взмахов сачка ***- численность на особей/м2
При проведении учета через 7 дней 28 мая перед вторым внесением препаратов было выявлено, что численность фитофагов на контроле возросла, а биологическая эффективность увеличилась во всех вариантах на 5-10 %, но сохранила те же тенденции, которые были выявлены на 3 день учета после обработки биоинсектицидами (таблица 5).
28 мая была проведена вторая обработка биоинсектицидами Биослип БВ, Биослип БТ и их смесью, а так же был применен химический способ борьбы смесью инсектицидов Алт-Альф, КЭ + Актара, ВДГ с нормами расхода 0,1 л/га и 0,05 кг/га. Озимая пшеница находились в этот момент в фазе цветения.
При учетах проведенных через 3 и 7 дней после применения препаратов были получены результаты, которые согласовывались с результатами первого применения биопрепаратов (таблиц 6).
На контроле через неделю численность одних фитофагов увеличилась вредной черепашки до 1,4 экз./ м2, злаковых тлей до 11,2 экз./растение и достигла ЭПВ (5-10 экз./ растение), пшеничного трипса до 15 экз./растение, численность других уменьшилась в связи с их биологией развития, так численность хлебных пилильщиков составляла 2,5 экз./ 100 взмахов сачка лет вредителя заканчивался, а численность пьявицы красногрудой снизилась до 0,7 экз./ растение, так как многие личинки окуклились (таблица 6). В варианте опыта с применением биоинсектицида Биослип БВ, ситуация поменялась нами бала отмечена относительно высокая биологическая эффективность в отношении не только злаковых тлей и вредной черепашки, но и пшеничного трипса – она составила через 7 дней после применения — 85, 73 и 65 % соответственно, наименьшая эффективность отмечена в отношении пьявицы красногрудой 42 и 39% на 3 и 7 день соответствено и по прежнему оставалась низкой в отношении хлебных пилильщиков 40-50 %.
В варианте опыта с применением биоинсектицида Биослип БТ, тенденция сохранилась и проявилась более контрастно, так как биологическая эффективность в отношении пьявицы красногрудой увеличилась до 87 %, а в отношении остальных видов фитофагов снизилась и колебалась в пределах 27-45 %.
В варианте опыта с применением баковой смеси биоинсектицидов Биослип БВ и Биослип БТ, биологическая эффективность против вредной черепашки, пшеничного трипса злаковых тлей и пьявицы красногрудой колебалась в пределах 50-62 %, наименьшая биологическая эффективность отмечена, как и при первом внесении препаратов, в отношении хлебных пилильщиков 30-35 % (таблица 7).
Таблица 6 — Изменение численности вредителей в агробиоценозе озимой пшеницы при применении биоинсектицидов и инсектицидов в фазу цветения (учебно-опытная станция, сорт Юка, 2019 г.)
| Объекты
наблюдения |
Число до обработки | Количество по суткам учетов после обработки | Биологическая эффективность, % | ||||
| 3 | 7 | 3 | 7 | ||||
| Биослип БВ (3 л/га) | |||||||
| Вредная черепашка*** | 0,15± 0,01 | 0,34± 0,01 | 0,38± 0,01 | 62 | 73 | ||
| Злаковые тли* | 0,8± 0,1 | 2,1± 0,1 | 1,7± 0,1 | 74 | 85 | ||
| Пшеничный трипс* | 4,2± 0,3 | 4,4± 0,3 | 5,3± 0,3 | 60 | 65 | ||
| Хлебные ** пилильщики | 3,4± 0,3 | 3,2± 0,2 | 1,3± 0,1 | 40 | 50 | ||
| Пьявица красногрудая* | 0,8± 0,01 | 1,1± 0,1 | 0,4± 0,01 | 42 | 39 | ||
| Биослип БТ (3 л/га) | |||||||
| Вредная черепашка*** | 0,25± 0,01 | 0,55± 0,02 | 0,77± 0,03 | 39 | 45 | ||
| Злаковые тли* | 2,0± 0,1 | 6,2± 0,4 | 8,1± 0,5 | 25 | 28 | ||
| Пшеничный трипс* | 6,0± 0,4 | 7,2± 0,5 | 9,0± 0,6 | 35 | 40 | ||
| Хлебные ** пилильщики | 3,5± 0,3 | 4,1± 0,3 | 1,8± 0,1 | 24 | 27 | ||
| Пьявица красногрудая* | 0,2± 0,01 | 0,4± 0,01 | 0,1± 0,01 | 81 | 87 | ||
| Биослип БВ + Биослип БТ (1,5 + 1,5 л/га) | |||||||
| Вредная черепашка*** | 0,23± 0,01 | 0,44± 0,01 | 0,56± 0,01 | 51 | 60 | ||
| Злаковые тли* | 1,6± 0,1 | 4,8± 0,3 | 5,6 | 42 | 50 | ||
| Пшеничный трипс* | 4,0± 0,3 | 5,7 | 6,8 | 48 | 55 | ||
| Хлебные ** пилильщики | 3,5± 0,3 | 3,8± 0,3 | 1,6± 0,1 | 30 | 35 | ||
| Пьявица красногрудая* | 0,5± 0,01 | 0,8± 0,01 | 0,3± 0,01 | 60 | 62 | ||
| Алт-Альф, КЭ + Актара, ВДГ (0,1 л/га + 0,05 кг/га) | |||||||
| Вредная черепашка*** | 0,7±0,05 | 0,01 | 0,01 | 99 | 99 | ||
| Злаковые тли* | 5,6± 0,5 | 0,4± 0,01 | 0,2± 0,01 | 95 | 98 | ||
| Пшеничный трипс* | 10 ± 0,8 | 0,3± 0,01 | 0,2± 0,01 | 97 | 99 | ||
| Хлебные ** пилильщики | 7,2±0,5 | 0,4± 0,01 | 0,1 | 93 | 97 | ||
| Пьявица красногрудая | 1,6± 0,2 | 0,1 | 0,0 | 97 | 98 | ||
| Контроль | |||||||
| Вредная черепашка*** | 0,7±0,05 | 0,9±0,07 | 1,4±0,1 | — | |||
| Злаковые тли* | 5,6± 0,5 | 8,2± 0,7 | 11,2± 1 | — | |||
| Пшеничный трипс* | 10 ± 0,8 | 11 ± 1 | 15 ± 1,2 | — | |||
| Хлебные ** пилильщики | 7,2±0,5 | 5,4±0,5 | 2,5±0,3 | — | |||
| Пьявица красногрудая* | 1,6± 0,2 | 1,9±0,2 | 0,7± 0,1 | — | |||
Примечание:*-численность на экз./колос (растение) **- численность на экз./ 10 взмахов сачка ***- численность на особей/м2
Таблица 7 — Изменение численности вредителей в агробиоценозе озимой пшеницы при применении биоинсектицидов и инсектицидов в фазу молочной спелости зерна (учебно-опытная станция СтГАУ, сорт Юка, 2019 г.)
| Объекты
наблюдения |
Число до обработки | Количество по суткам учетов после обработки | Биологическая эффективность, % | |||||
| 3 | 7 | 3 | 7 | |||||
| Биослип БВ (3 л/га) | ||||||||
| Вредная черепашка*** | 0,38± 0,01 | 0,54± 0,01 | 0,65± 0,01 | 73 | 76 | |||
| Злаковые тли* | 1,7± 0,1 | 2,1± 0,1 | 2,5± 0,1 | 85 | 87 | |||
| Пшеничный трипс* | 5,3± 0,3 | 6,7±0,5 | 7,8±0,5 | 65 | 69 | |||
| Хлебные ** пилильщики | 1,3± 0,1 | 1,1± 0,1 | 0,3± 0,01 | 50 | 73 | |||
| Пьявица красногрудая* | 0,4± 0,01 | 0,3± 0,01 | 0,2± 0,01 | 39 | 25 | |||
| Биослип БТ (3 л/га) | ||||||||
| Вредная черепашка*** | 0,77± 0,03 | 1,1± 0,1 | 1,57± 0,1 | 45 | 42 | |||
| Злаковые тли* | 8,1± 0,5 | 10,2± 1 | 13,12± 1 | 28 | 32 | |||
| Пшеничный трипс* | 9,0± 0,6 | 11,4± 1 | 14,25± 1 | 40 | 43 | |||
| Хлебные ** пилильщики | 1,8± 0,1 | 1,5± 0,1 | 0,77± 0,01 | 27 | 30 | |||
| Пьявица красногрудая* | 0,1± 0,01 | 0,1 | 0,02 | 87 | 90 | |||
| Биослип БВ + Биослип БТ (1,5 + 1,5 л/га) | ||||||||
| Вредная черепашка*** | 0,56± 0,01 | 0,8± 0,04 | 1,22± 0,1 | 60 | 55 | |||
| Злаковые тли* | 5,6 | 7,1±0,5 | 8,3±0,5 | 50 | 57 | |||
| Пшеничный трипс* | 6,8 | 8,6±0,5 | 10,0± 1 | 55 | 60 | |||
| Хлебные ** пилильщики | 1,6± 0,1 | 1,4± 0,1 | 0,7± 0,03 | 35 | 40 | |||
| Пьявица красногрудая* | 0,3± 0,01 | 0,2± 0,01 | 0,1 | 62 | 60 | |||
| Алт-Альф, КЭ + Актара, ВДГ (0,1 л/га + 0,05 кг/га) | ||||||||
| 10 сутки | 14 сутки | |||||||
| Вредная черепашка*** | 0,01 | 0,0 | 0,0 | 99 | 100 | |||
| Злаковые тли* | 0,2± 0,01 | 0,2± 0,01 | 0,2± 0,01 | 98 | 98 | |||
| Пшеничный трипс* | 0,2± 0,01 | 0,2± 0,01 | 0,2± 0,01 | 98 | 99 | |||
| Хлебные ** пилильщики | 0,1 | 0,1 | 0,0 | 98 | 100 | |||
| Пьявица красногрудая | 0,1 | 0,0 | 0,0 | 97 | 100 | |||
| Контроль | ||||||||
| Вредная черепашка*** | 1,4±0,1 | 2,0±0,2 | 2,7±0,2 | — | ||||
| Злаковые тли* | 11,2± 1 | 14,1± 2 | 19,3± 2 | — | ||||
| Пшеничный трипс* | 15 ± 1,2 | 19 ± 2 | 25± 3 | — | ||||
| Хлебные ** пилильщики | 2,5±0,3 | 2,1±0,5 | 1,1±0,4 | — | ||||
| Пьявица красногрудая* | 0,7± 0,01 | 0,5±0,01 | 0,2± 0,01 | — | ||||
Примечание:*-численность на экз./колос (растение) **- численность на экз./ 10 взмахов сачка ***- численность на особей/м2
В варианте, где применяли баковую смесь инсектицидов Алт-Альф, КЭ + Актара, ВДГ с нормами расхода 0,1 л/га и 0,05 кг/га учёты на 3 и 7 день после обработки показали, что биологическая эффективность против вредителей составила 93-99 %.
После проведения учетов 4 июня была проведена третья обработка биоинсектицидами Биослип БВ, Биослип БТ и их смесью, озимая пшеница находились в фазе молочной спелости зерна.
В варианте без обработки инсектицидами численность вредной черепашки, злаковых тлей и пшеничного трипса продолжала увеличиваться, а численность имаго хлебных пилильщиков, в связи с их естественной гибелью после откладки яиц и пьявицы красногрудой, в связи с окукливанием личинок, снизилась (таблица 7).
В варианте опыта с применением биоинсектицида Биослип БВ, биологическая эффективность продолжала увеличиваться в отношении злаковых тлей, вредной черепашки и пшеничного трипса через 7 дней после применения она составила – 87, 76 и 69 % соответственно, изменилась ситуация в отношении личинок хлебных пилильщиков биологическая эффективность в отношении этого вида к третьей обработки увеличилась с 50 до 73 %, и наименьшая биологическая эффективность отмечена в отношении пьявицы красногрудой 39 и 25 % на 3 и 7 день соответствено. В варианте опыта с применением биоинсектицида Биослип БТ, тенденции так же сохранились, как и после первой и второй обработки, так биологическаяэффективность в отношении пьявицы красногрудой достигла до 90 %, и стала сопоставима с химическими препаратами, а в отношении остальных видов фитофагов она колебалась в пределах 27-45 %. В варианте опыта с применением баковой смеси биоинсектицидов Биослип БВ и Биослип БТ, биологическая эффективность против вредной черепашки, пшеничного трипса злаковых тлей и пьявицы красногрудой колебалась в пределах 50-62 %, наименьшая биологическая эффективность отмечена, как и при первом внесении препаратов, в отношении хлебных пилильщиков 35-40 %. В варианте, где применяли баковую смесь инсектицидов Алт-Альф, КЭ + Актара, ВДГ учёты на 10 и 14 день после обработки показали, что биологическая эффективность против вредителей составила 97-100 % .
4.4 Определение токсичности биоинсектицидов для различных групп энтомофагов озимой пшеницы
Число видов членистоногих, обнаруживаемых на полях с любой сельскохозяйственной культурой и в любых географических зонах, обычно достигает нескольких сотен. На первом месте по числу видов чаще всего оказываются энтомофаги(паразиты и хищники), на втором– фитофаги, а на третьем – детритофаги. Примерно столько же, как и детритофагов, вылавливается членистоногих, не имеющих прямого отношения к агроэкосистеме. Очень незначительно число видов антофилов. После внесения органических удобрений на поле могут быть найдены некоторые виды копрофагов, в основном, жуков и двукрылых.
Монокультура существенно снижает видовое разнообразие фитофагов на поле. Число их видов заметно увеличивается в том случае, если на поле есть сорняки. Набор паразитических видов, хотя и связан с комплексом фитофагов, но значительно меньше зависит от сельскохозяйственной культуры. Хищники-генералисты почти всегда одни и те же на всех культурах, хотя разный микроклимат на полях и даже в пределах одного поля может влиять на количественное соотношение видов.
Комплексы фитофагов хорошо изучены и подробно описаны в учебниках сельскохозяйственной энтомологии. Значение детритофагов как дополнительных жертв энтомофагов может быть очень большим, потому что они встречаются на поле даже при отсутствии там наземной растительности и, соответственно, и фитофагов. Однако роль этой экологической группы в привлечении энтомофагов на поле и поддержании естественного баланса совершенно неизвестна. Здесь мы остановимся только на основных группах энтомофагов и акарифагов, наиболее типичных для агроэкосистем и обеспечивающих экологическое равновесие в пределах комплекса членистоногих.
Особое внимание следует уделить неспециализированным энтомофагам, так как именно они играют ключевую роль в регуляции численности вредителей. Некоторая специализация– преимущественное питание тлями– имеет место у афидофагов, однако большинство из них способны использовать и другую пищу, например, яйца насекомых. Конечно, видовой состав членистоногих зависит от географического района.
Несмотря на мнение многих исследователей о том, что регуляторная роль хищных и паразитических насекомых в агробиоценозах значительно ослаблена, по сравнению с природными экосистемами, проведено много исследований по выявлению видов энтомофагов, способных влиять на численность фитофагов в посевах озимой пшеницы. В результате этого было, выявлено свыше 90 видов многоядных хищников (жужелицы, стафилиниды, кокцинеллицы, хризопиды, мухи-журчалки, аэлотрипиды, клопы набиды, пауки и др.), питающихся фитофагами этой культуры.
При этом была установлена предпочтительность питания хищников определёнными фазами развития вредителей. Так яйцами фитофагов питаются несколько видов пауков, ряд видов жуков из семейств Coccinellidae, Anthicidae и Carabidae и других хищных насекомых. Имаго и личинок клопов, трипсов и тлей охотно поедают жужелицы родов Pterostichus, Dolichus, Bembidion, Harpalus, Calosoma и др., пауки семейств Thomisidae Lycosidae, жуки семейств Coccinellidae, Anthicidae, Staphylinidae и муравьи.
Оказалось, что наиболее активными многоядными хищниками являются жуки сем. Coccinellidae и Carabidae, мухи сем. Syrphidae, пауки семейств Thomisidae и Lycosidae, но их эффективность сильно варьирует по годам и агроклиматическим зонам страны.
Помимо хищных членистоногих, значительное внимание исследователей было уделено изучению видового состава, биологии и эффективности паразитов наиболее значимых вредителей озимой пшеницы, особенно вредной черепашки. Было установлено, что в имаго клопа вредной черепашки паразитирует 4 вида мух фазий. Но как показали исследования, данные паразиты слабо ограничивают численность клопа в районах с благоприятными для его развития климатическими условиями, несмотря на то, что серая (Alophora subcoleoptrata L.) и пестрая (Phasia (Ectophasia) crassipennis F.), фазии являются специализированными паразитами клопов щитников.
Другими паразитами вредной черепашки являются яйцееды. В результате проведённых исследований было выявлено 13 видов паразитов из 7 родов. Наиболее распространёнными и многочисленными являются представители р. Trissolcus и Telenomus. Установлено, что эффективность яйцеедов сильно варьирует в зависимости от климатических условий региона. Так, в засушливых условиях Ростовской, Саратовской и Воронежской областей отмечена низкая эффективность яйцеедов в снижении численности клопа вредной черепашки. Напротив, в таких регионах, как предгорья Краснодарского края, Ставропольская возвышенность, в Азербайджане, Украине, Казахстане и Болгарии теленомины в отдельные годы значительно снижали численность вредной черепашки.
С целью повышения эффективности природных энтомофагов в снижении численности фитофагов в посевах озимой пшеницы, были проведены исследования по использованию паразитов и хищников методом сезонной колонизации. В производственных опытах, выполненных в начале 80 гг. прошлого столетия в Харьковской и Воронежской областях и в Краснодарском крае, было установлено, что эффективность теленомин, при таком способе борьбы с вредной черепашкой не превышает 20 %. Также малоэффективными оказались выпуски в посевы зерновых культур таких хищников, как златоглазки и кокцинеллиды.
Дальнейшими исследованиями было показано, что более перспективно в борьбе с вредной черепашкой использование природных популяций теленомин путём повышение их эффективности за счёт рационального размещения полей пшеницы в структуре сельскохозяйственных угодий, разнообразия выращиваемых культур, граничащих с полями пшеницы посевов нектароносов, наличие естественных стаций с разнообразием цветущих растений весной.
Во ВНИИБЗР был разработан метод увеличения зимующего запаса телемин, основанный на их развитии на площадках с имаго и кладками яиц клопа графазомы, размещённых на пропашных культурах (кукуруза, соя, подсолнечник), граничащих с посевами озимой пшеницы, куда они мигрируют во второй половине сезона после прекращения откладки яиц вредной черепашкой.
Важным фактором сохранения природных энтомофагов являются лесополосы, влияющие с одной стороны на микроклимат граничащих с ними полей озимой пшеницы, а с другой являющиеся местообитанием многих видов фитофагов и эитомофагов благодаря произрастанию в них и на их опушках различных цветущих растений-нектароносов.
Пространственное распределение, т.е. характер расселения организмов на полях экосистем или на полях сельскохозяйственных культур, один из важнейших показателей состояния популяций и сообществ в их взаимодействии со средой обитания и между собой. Характер распределения насекомых на поле зависит от многих факторов (образа жизни насекомых, способа питания, наличия развитого летательного аппарата, предшествующей культуры и др.), но главным образом он зависит от особенностей вида и размера насекомого.
В полевых исследованиях определение пространственного расселения видов членистоногих весьма трудоемкий процесс, непосредственно связанный с учетами их численности. Мониторинг численности отдельных видов, как правило, проводится методом случайной выборки, а число отбираемых проб зависит от размеров обследуемой площади, и требует больших временных затрат. В связи с этим простым и удобным методом описания пространственного распределения насекомых является метод сравнения эмпирических серий с теоретическими статистическими распределениями (ТСР). Существует много теоретических распределений, которые в принципе можно использовать для формализации эмпирических серий данных (Пуассона, нормальное распределение, гамма, и др.). Однако наиболее информативно значимые результаты дают ТСР базирующиеся на моделях расселения. Параметры таких теоретических статистических распределений интерпретируемы и могут быть оценены прямыми экспериментальными исследованиями).
При изучении консортов первого и второго порядка в системе триотрофа озимой пшеницы, мы заметили, что плотность поселения особей к середине поля снижается. И мы задались вопросом насколько это носит закономерный характер.
Исходя из проведенных обследований, мы рассчитали среднюю численность популяции изучаемых видов насекомых для различных точек агробиоценоза озимой пшеницы, исходя из собранного материала. Результаты обобщения представлены в таблице 8 и отражают общие тенденции распределения видов членистоногих на посеве озимой пшеницы.
Самая большая численность, как вредителей, так и их энтомофагов, как правило, отмечается на краях поля. По мере удаления от краев их численность уменьшается более или менее резко, в соответствии с образом жизни и поведением того или иного вида насекомого.
Вредная черепашка — крупный клоп и имеет хорошо развитый летательный аппарат, легко передвигается по всему полю озимой пшеницы. К середине поля его численность снижается только в 2 раза. Паразиты яиц (теленомины) мелкие насекомые и нуждаются в дополнительном питании на нектароносной растительности, В связи с этим максимальная их численность наблюдается по краям поля, а к середине она уменьшается в три раза, то есть для их распределения характерен ярко выраженных краевой эффект.
Распределение по полю мух фазий, которые являются хорошо и активно летающими насекомыми, случайно распределяющимися по полям озимой пшеницы, и их численность снижается к середине поля пропорционально численности черепашки. Кроме того, личинки фазий дополнительно разносятся перезимовавшими клопами.
Таблица 8 – Численность фитофагов и энтомофагов в агробиоценозе озимой пшеницы
| Вид насекомого | Средняя численность на 1 м в зависимости от удаленности от края поля (м) | |||||
| 50 | 100 | 200 | 300 | 400 | 500 | |
| Вредная черепашка | 5,8±0,5 | 4,2±0,4 | 3,6±0,3 | 3,0±0,3 | 2,8±0,3 | 2,6±0,3 |
| Теленомины | 9,1±1,2 | 8,3±1,1 | 7,2±1,1 | 5,8±0,9 | 4,9±0,8 | 3,4±0,7 |
| Фазии | 3,1±0,3 | 2,8±0,3 | 2,0±0,2 | 2,2±0,2 | 1,8±0,2 | 1,5±0,2 |
| Злаковые тли | 1298,4± 87,3 | 1118,7± 81,2 | 1004,3± 74,2 | 934,8± 65,3 | 812,4± 59,5 | 746,7± 51,9 |
| Кокцинеллиды | 20,2±3,2 | 18,3±3,0 | 16,0±2,8 | 14,4±2,4 | 13,2±2,0 | 11,8±1,8 |
| Сирфиды | 8,4±1,7 | 6,0±1,5 | 5,2±1,1 | 4,4±0,9 | 3,8±0,7 | 3,2±0,6 |
| Златоглазки | 3,1±0,5 | 2,7±0,4 | 2,5±0,3 | 2,1±0,2 | 1,8±0,2 | 1,4±0,2 |
| Афидииды | 12,2±1,7 | 10,1±1,6 | 8,5±1,4 | 6,2±1,1 | 5,3±0,8 | 3,9±0,6 |
| Пшеничный трипс | 42,0±3,5 | 37,3±3,3 | 33,4±3,1 | 28,2±2,6 | 24,3±2,4 | 20,5±1,8 |
| Полосатый трипс | 0,9±0,01 | 0,7±0,01 | 0,6±0,01 | 0,5±0,01 | 0,5±0,01 | 0,4±0,01 |
| Обыкновенный хлебный пилильщик | 5,1±1,2 | 4,5±0,5 | 4,2±0,2 | 3,7±0,2 | 3,4±0,2 | 3,0±0,2 |
| Коллирия | 2,8±0,2 | 2,6±0,2 | 2,3±0,1 | 1,9±0,1 | 1,7±0,1 | 1,5±0,1 |
| Всего | 1411,1±
95,6 |
1216,2± 84,8 | 1089,8± 78,9 | 1007,2± 71,7 | 875,9± 65,6 | 799,9± 58,9 |
Злаковые тли по характеру распределения в агробиоценозах относится к агрегативным видам, т.е. к видам, образующим колонии. По нашим данным средняя численность злаковых тлей к середине поля озимой пшеницы сокращается в 1,7 раза, что составляло 42,5% от численности тлей в среднем по агробиоценозу. На краевых полосах поля (50-100 метров от края) численность данного фитофага составляет в среднем 1298,4 экз./ м , к центру поля численность снижается до 746,7 экз./ м2. Из энтомофагов только численность кокцинеллид, снижается пропорционально снижению численности жертвы в агробиоценозе — в 1,7 раза. Снижение численности других её энтомофагов (сирфид, хризопид и афидииды), нуждающихся в дополнительном, снижалось в большей мере. Так количество имаго и личинок сирфид и златоглазок снижалось в 2-2,5 раза, афидиид — в 3,1 раза.
При выявлении пространственного распределения в агробиоценозе озимой пшеницы пшеничного трипса, нами отмечалось снижение численности его популяции к середине поля в 2 раза, так в среднем за 5 лет на краю поля его численность составила 42,0±3,5 экз/колос, в середине поля — 20,5±1,8 экз/колос.
Снижение численности популяции его специализированного хищника — полосатого трипса к середине агробиоценоза составило 2,2 раза, то есть она коррелируется с численностью его жертвы.
Численность обыкновенного хлебного пилильщика к центру поля сокращается в 1,7 раза, а его паразита коллирии — в 1,8 раза.
Из приведенных данных можно сделать вывод, что в среднем численность фитофагов и их энтомофагов к середине поля сокращается в 1,7-3,0 раза.
Для более объективной оценки характера распределения видов насекомых на полях озимой пшеницы провели статистическую обработку полученных данных с помощью градиента анализа с использованием следующих величин:
Q — индекс меры «неупорядоченности» матрицы сходства;
Qrand — индекс ожидаемое значение Q для «полностью неупорядоченной»
матрицы сходства;
Q/Qmax — соотношение индексов меры сходства;
P< — оценка статистической значимости градиента; Так, при P < 0,01, наблюдаемая упорядоченность матрицы неслучайна с вероятностью более 99%. Если же Р > 0,10, то отличия исследуемой матрицы от неупорядоченной недостоверны.
Мы привлекли дополнительную оценку степени упорядоченности матрицы, величину коэффициента корреляции (R). Высокое, статистически значимое отрицательное значение R указывает на существование достоверного градиента видовой структуры. Бета-разнообразие, характеризует степень различий между разными выборками.
S — качественный индекс меры сходства структурной гетерогенности (мозаичности);
B — количественный индекс меры сходства — «дистанция между пробами», взятый с обратным знаком (степень градиентности).
Di/D2 — мера дискретности градиента.
Из приведенных в таблице 9 значений видим, что показатели P и R ежегодно подтверждают наличие статистически значимого градиента изменения численности видов, наблюдаемого по направлению от края к середине поля. Эти показатели свидетельствуют о том, что численность популяций насекомых постепенно уменьшается по напралению к центру, что подтверждается количественным индексом меры сходства (B) и мерой дискретности градиента (Di/D2).
На основании экспериментальных данных, приведенных в таблице 8 были рассчитаны аналитические уравнения регрессий, отражающие закономерности изменения численной структуры популяций насекомых в различных точках агробиоценоза озимой пшеницы (таблица 9 ).
Таблица 9 – Аналитические уравнения регрессий изменения численности консументов первого и второго порядка в различных точках агробиоценоза озимой пшеницы
| Вид насекомого | Уравнения регрессии | Достоверность апроксимации R2 |
| Вредная черепашка | y = 0,1536x2 — 1,6693x + 7,18 | 0,9843 |
| Теленомины | y = -0,0482x2 — 0,8082x + 10,01 | 0,9967 |
| Фазии | y = 0,0286x2 — 0,5086x + 3,58 | 0,9155 |
| Злаковые тли | y = 1,75×2 — 122,45x + 1406,2 | 0,9678 |
| Кокцинеллиды | y = 0,1232x2 — 2,5454x + 22,69 | 0,9978 |
| Сирфиды | y = 0,175x2 — 2,1793x + 10,14 | 0,9775 |
| Златоглазки | y = -0,0071x2 — 0,2814x + 3,36 | 0,9953 |
| Афидииды | y = 0,1125x2 — 2,4504x + 14,57 | 0,9951 |
| Пшеничный трипс | y = 0,0804x2 — 4,8968x + 46,87 | 0,9987 |
| Полосатый трипс | y = 0,0161x2 — 0,2039x + 1,07 | 0,9746 |
| Обыкновенный хлебный пилильщик | y = 0,0179x2 — 0,5336x + 5,58 | 0,9949 |
| Коллирия | y = 0,0071x2 — 0,3243x + 3,16 | 0,9889 |
Полученные уравнения можно использовать для оптимизации учетов при проведении на больших посевах озимой пшеницы исследований по рас счету средней плотности популяции в определенном агробиоценозе, с поправкой на уменьшение численности насекомых от края поля к его середине.
В целом статистический анализ данных по распределению доминантных видов членистоногих на полях озимой пшеницы, свидетельствует о том, что крупные поля озимой пшеницы, наиболее уязвимы в середине поля, так как там соотношение фитофагов и их энтомофагов ниже, чем на краевых полосах поля. При благоприятных погодных условиях именно там может произойти резкое увеличение численности фитофагов и снижение регулирующей роли природных энтомофагов даже при соблюдении организационных и технологических приемов возделывания. Особенно это актуально для фитофагов имеющих ярко выраженный тип отрицательного биноминального распределения в агробиоценозе, что характерно для злаковых тлей и пшеничного трипса.
В связи с этим, при принятии решения по обработке крупных полей, необходимо детальное обследование посевов с целью выявления участков, где численность фитофагов превышает экономический порог вредоносности, и природные энтомофаги не могут оказать существенного влияния на снижение их численности. Для большей эффективности яйцеедов и афидофагов можно рекомендовать оптимизацию площадей полей, а при большой площади поля делать полосные посевы нектароносов.
Результаты исследований показывают, что в Центральном Предкавказье в зоне неустойчивого увлажнения природные ресурсы консортов второго порядка (энтомофагов) в консорции озимой пшеницы многочисленны и являются постоянно действующим биотическим фактором, ограничивающим рост популяций консортов первого порядка. Одним из путей целенаправленного использования этих ресурсов в интегрированной защите озимой пшеницы является определение уровней эффективности природных популяций энтомофагов и на этой основе внесение корректив в тактику защитных мероприятий.
В связи с этим в 2019 в посевах озимой пшеницы на учебно-опытной станции Ставропольского ГАУ мы проводили исследования по эффективности природных энтомофагов в садках размером 3 м2 (1х3 м), размещённых вдоль рядков озимой пшеницы в трехкратной повторности. В период вегетации пшеницы на VIII этапе органогенеза выпускали расчетное количество клопов вредной черепашки (6 или 8 имаго), среди которых были клопы зараженные фазиями, их количество от общего числа выпускаемых клопов указано через дробь в таблице 10,. Зараженность яиц вредной черепашкой теленоминами производили после откладки яиц, запуская паразитов в садок и, при заражении нужного числа яиц, паразитов из садка удаляли. В качестве контроля использовали химическую обработку, которую проводили баковой смесью инсектицидов Алт-Альф, КЭ + Актара, ВДГ в норме применения 0,1 + 0,05 л/га против вредной черепашки при достижении имаго численности 2 экз./м2 и 0,1 л/га против трипсов и тлей.
Аналогичным образом были проведены исследования по изучению эффективности афидофагов и полосатого трипса.
Наши исследования показали, что если зараженность перезимовавших клопов мухами фазиями при численности 2 экз/м2 составляет 33 %, а зараженность яиц вредной черепашки теленоминами — 40%, то необходимость химической обработки отпадает (таблица 10).
Таблица 10 -Эффективность паразитов вредной черепашки в агробиоценозе озимой пшеницы
| Численность вредной черепашки, экз/м2 | Зараженность клопов
фазиями, % |
Зараженность яиц вредной черепашки теленоми- нами,% | Урожай, т/ га | Содержание
клейковины, % |
ИДК | Стекловид-
ность, % |
| Химическая обработка | 4,56 | 26,7 | 85 | 73 | ||
| 2 (6/1) | 16 | 20 | 4,14 | 22,7 | 95 | 65 |
| 2 (6/2) | 33 | 40 | 4,14 | 26,4 | 90 | 70 |
| 2,6 (8/2) | 25 | 20 | 4,14 | 21,6 | 95 | 63 |
| 2,6 (8/3) | 37 | 40 | 4,24 | 22,3 | 90 | 65 |
Исследования эффективности афидофагов злаковых тлей показали, что химическую обработку без риска для качества и количества урожая можно отменить при соотношении хищников и злаковых тлей 1 : 30-40 без учета количества паразитированных тлей. Комплекс хищников сдерживает вредителей на хозяйственно неощутимом уровне при соотношении хищник- жертва на озимой пшенице 1:40-50 при одновременном заражении тлей паразитами не ниже 30% популяции (таблица 11).
Таблица 11 — Эффективность афидофагов злаковых тлей в агробиоценозе
озимой пшеницы
| Численность вредной черепашки, экз/м2 | Зараженность клопов
фазиями, % |
Зараженность яиц вредной черепашки теленоми- нами,% | Урожай, т/ га | Содержание
клейковины, % |
ИДК | Стекловид-
ность, % |
| Химическая обработка | 4,56 | 27,2 | 85 | 74 | ||
| 1 | 30-40 | 15 | 4,14 | 70 | 95 | 65 |
| 1 | 30-40 | 30 | 4,14 | 71 | 90 | 70 |
| 1 | 40-50 | 15 | 4,24 | 67 | 95 | 63 |
| 1 | 40-50 | 30 | 4,24 | 68 | 90 | 65 |
| 1 | 50-60 | 15 | 4,14 | 65 | ||
| 1 | 50-60 | 30 | 4,24 | 66 | ||
Из полученных результатов видно, что при соотношении полосатого и пшеничного трипсов 1:40-50 можно гарантированно отменять химическую обработку, при сотношении 1:50-60 урожай в количественном выражении снижается незначительно по сравнению с контролем, но содержание клейковины уменьшается на 3,6% (таблицы 12).
Таблица 12 – Эффективность полосатого трипса — хищника пшеничного трипса в агробиоценозе озимой пшеницы
| Соотношение полосатого
и пшеничного трипсов |
Урожайность,
т/га |
Содержание клейковины, % | ИДК | Стекловидность, % |
| Химическая обработка | 4,56 | 26,8 | 85 | 77 |
| 1 : 40-50 | 4,14 | 26,5 | 88 | 74 |
| 1 : 50-60 | 4,14 | 23,2 | 92 | 70 |
| 1 : 60-70 | 4,24 | 21,3 | 95 | 63 |
Проведенные исследования позволяют нам рекомендовать отмену химических обработок посевов озимой пшеницы:
— против вредной черепашки при численности имаго 2 экз./м2, их 25 % зараженности мухами фазиями и 40% заражённости яиц вредной черепашки теленоминами;
— против злаковых тлей при соотношении хищник : жертва 1:30-1:40 и при соотношении 1:40-50, если зараженность тлей паразитами не ниже 30%; — против пшеничного трипса при соотношении хищник : жертва 1:40-50. Применение данных рекомендаций в производственных условиях позволит сократить применение химических средств защиты растений на посевах озимой пшеницы, сохранить природных энтомофагов и уменьшить загрязнение окружающей среды от пестицидов.
4.5. Сравнительная оценка вариантов возможных решений поставленных задач
Про проведенным учетам нами была рассчитана средняя биологическая эффективность изучаемых вариантов опыта рисунок 17.
Полученные результаты исследований показали, что биоинсектицид Биослип БВ с нормой расхода 3 л/га, наиболее эффективно подавляет в посевах агроценоза озимой пшеницы злаковых тлей. Его средняя биологическая эффективность в отношении данного вредителя составила 78,3 %, а после третьей обработки через 7 дней она составляла 87 %. Так же данный препарат обладает средней биологической эффективностью в отношении вредной черепашки и пшеничного трипса, 60,3-68,7 %. Биоинсектицид Биослип БТ, как показали результаты опыта хорошо подходит для защиты посевов от пьявицы красногрудой, так ка его средняя биологическая эффективность составила 84,2 %, что сопоставима с химической защитой, в отношении других видов вредителей его эффективность низкая и колебалась в пределах 27,2-44,2 %. Применение баковой смеси в половинных нормах расхода данных препаратов не целесообразно, так как эффективность данной смеси колебалась в пределах 23,3-40,7 %, что не достаточно для подавления роста популяции фитофагов в посевах озимой пшеницы и ограничения их вредоносности.
8 июля 2019 года провели уборку урожая озимой пшеницы методом прямого комбайнирования с измельчением соломы комбайном New Holland. Уборочная влажность зерна 9,5 %.
Результаты уборки по влиянию биоинсектицидов и инсектицидов на урожайность озимой пшеницы сорта Юка в 2019 году в ФГБОУ ВО СтГАУ учебно-опытная станция представлены в таблице 13.
Итак, как показали результаты уборки 2019 года видим, что вредоносность фитофагов в посевах озимой пшеницы велика, так как урожайность на контроле ниже на 0,556 т/га по сравнению с химической обработкой в фазу цветения.
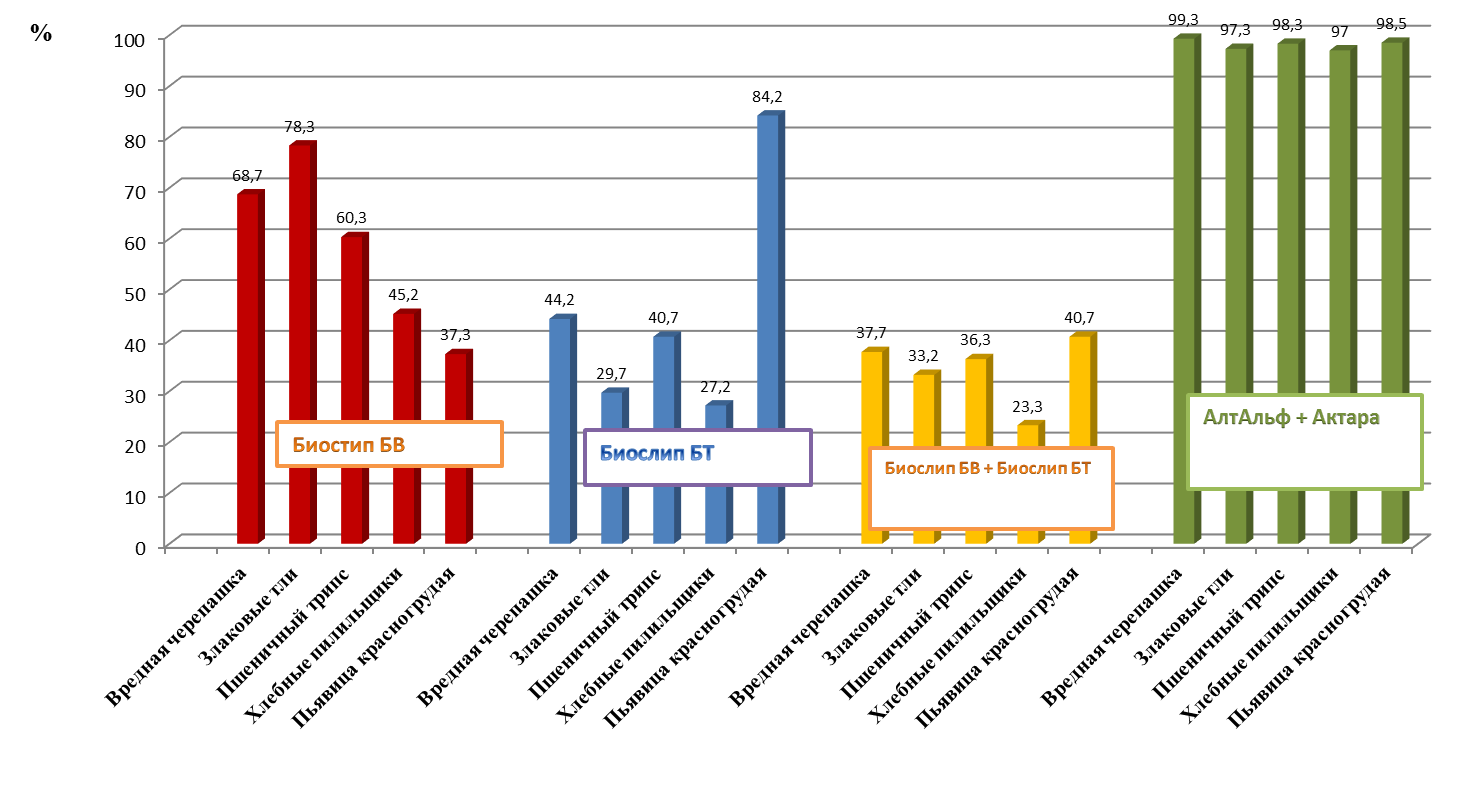
Рисунок 17 – Биологическая эффективность биоинсектицидов и инсектицидов против вредителей в агробиоценозе озимой пшеницы (учебно-опытная станция СтГАУ, сорт Юка 2019 г.)
Из полученных результатов уборки видно, что наименьшая разница с хозяйственным этолоном в варианте с трехкратным применением Биослипа БВ 0,09 т/га, на в вариантах с трехкратным применением препарата Биослип БТ и смеси с половинными его с препаратом Биослип БВ снижение в урожайности составило 0,32-0,42 т/га соответственно.

Рисунок 18 — Уборка опытов озимой пшеницы сорта Юка в 2019 году в ФГБОУ ВО СтГАУ учебно-опытная станция.
По полученной урожайности мы видим, что количество урожая напрямую зависит от эффективности применяемого биоинсектицида, так как наиболее эффективным в период обработок был Биослип БВ, против клопа вредной черепашки, злаковых тлей, пшеничного трипса и хлебных пилильщиков, только в отношении пьявицы красногрудой его эффективность была низкой.
Таблица 13 – Влияние биоинсектицидов на урожайность озимой пшеницы сорта Юка
(учебно-опытная станция ФГБОУ ВО Ставропольский ГАУ, 2019 г.)
| № | Вариант опыта | Урожайность, т/га | Прибавка по отношению к контролю, т/га | Разница с хозяйственном эталоном, т/га |
| 1 | Контроль (без обработки) | 4,0 | — | — |
| 2 | Биослип БВ | 4,47 | 0,47 | — 0,09 |
| 3 | Биослип ВТ | 4,24 | 0,24 | — 0,32 |
| 4 | Биослип БВ + Биослип ВТ | 4,14 | 0,14 | — 0,42 |
| 5 | Алт-Альф, КЭ + Актара, ВДГ | 4,56 | 0,56 | — |
Итак, при разработке системы биологической защиты озимой пшеницы в весенне-летний период вегетации от вредителей, на основе полученных результатов, следует, что при защите от пьявицы красногрудой необходимо использовать биоинсектицид Биослип БТ с нормой расхода 3 л/га, в дальнейшем вторую обработку провести через 7-10 дней баковой смесью биоинсектицидов Биослип БВ и Биослип БТ с нормами расхода 3 + 3 л/га, так как в условиях Центрального Предкавказья отмечается нарастание численности комплекса вредителей озимой пшеницы (клоп вредная черепашка, злаковые тли, пшеничный трипс, хлебные пилильщики), и третью обработку проводить через 7-10 дней биоинсектицидом Биослип БВ с нормой расхода 3 л/га.
5 РАСЧЕТ ЭКОНОМИЧЕСКОЙ ЭФФЕКТИВНОСТИ БИОЛОГИЧЕСКОЙ ЗАЩИТЫ ОЗИМОЙ ПШЕНИЦЫ ОТ ВРЕДИТЕЛЕЙ
Одним из главных путей подъема сельского хозяйства и удовлетворения возросших потребностей страны в сельскохозяйственной продукции является резкое повышение урожайности всех культур и увеличение выхода продукции с каждого гектара при наименьших затратах труда и средств. Рост продукции сельского хозяйства должен опережать растущий спрос на нее. Экономическая эффективность применения препаратов биологического происхождения в посевах озимой пшеницы представлена в таблице 14.
Таблица 14 – Экономическая эффективность применения биоинсектицидов в посевах озимой пшеницы сорта Юка
(учебно-опытная станция ФГБОУ ВО Ставропольский ГАУ, 2019 г.)
| Показатели | Варианты опыта | |||||
| Контроль | Биослип БВ | Биослип ВТ | Биослип БВ + Биослип ВТ | Алт-Альф + Актара | ||
| 1 | Урожайность с 1 га, т | 4,00 | 4,47 | 4,24 | 4,14 | 4,56 |
| 2 | Цена реализации 1 т, руб. | 10000 | 10000 | 10000 | 10000 | 10000 |
| 3 | Денежная выручка с 1 га, руб. | 40000 | 44700 | 42400 | 41400 | 45600 |
| 4 | Затраты труда на 1 га, чел.-ч. | 9,3 | 12,9 | 12,8 | 12,6 | 10,7 |
| 5 | Затраты труда на 1 т, чел.-ч. | 2,3 | 2,9 | 3,0 | 3,0 | 2,3 |
| 6 | Производственные затраты на 1 га, руб. | 25071,0 | 31546,9 | 31494,6 | 31294,7 | 26795,0 |
| 7 | Себестоимость 1 т продукции, руб. | 6267,7 | 7057,5 | 7428,0 | 7559,1 | 5876,1 |
| 8 | Прибыль на 1 га, руб. | 14929,0 | 13153,1 | 10905,4 | 10105,3 | 18805,0 |
| 9 | Уровень рентабельности,% | 59,5 | 41,7 | 34,6 | 32,3 | 70,2 |
Большие потери зерна озимой пшеницы связаны с повреждением ее фитофагами. Нередко ее продуктивность снижается на 10-20 % и более. В результате повреждения ухудшается качество зерна.
Многими исследователями показано, что, помимо внешних признаков повреждения, питание вредной черепашки на зерновках пшеницы приводит к ухудшению хлебопекарных качеств муки вследствие действия протеиназ вредителя на клейковинные белки эндосперма. Протеолитические ферменты вредителя при замешивании теста растворяются в воде и быстро расщепляют клейковинообразующие белки глиадины и глютенины.
Согласно данным таблицы 14, наибольшая общая прибыль получена в варианте с химическими препаратами, что объясняется высоким показателем урожайности в этом варианте опыта. Она составила 18 8105,0 тысяч рублей, что на 3 876,0 тысяч рублей больше, чем на контроле. Кроме того, это в среднем на 5 652,0-8 700,0 тыс. рублей больше, чем в вариантах с биопрепаратами.
Анализируя экономические показатели производства озимой пшеницы, мы обратили внимание на производственные затраты на защитные мероприятия в расчете на 1 га. Двухкратная обработка биоинсектицидами повысила производственных затрат до уровня 31 294,7-31 546,9 тыс. руб. на 1 га по сравнению 26 795,0 тыс. руб. в варианте с химическими препаратами и 25 ,741,0 тыс. руб. – на контроле.
Уровень рентабельности в варианте с химической обработкой практически 2 раза превышал показатели в вариантах с биологическими обработками.
Применение инсектицидов биологического происхождения повышает себестоимость производства 1 т продукции в среднем на 1 181,0-1 683,0 тыс. руб. по сравнению с пестицидными обработками и на 789,8-1 291,0 тыс. руб. по сравнению с контролем.
ЗАКЛЮЧЕНИЕ
В результате исследований, выполненных в Центральном Предкавказье, выявлен вредоносный комплекс доминантных видов фитофагов, повреждающих озимую пшеницу в весенне-летний период вегетации. Данный комплекс представлен вредной черепашкой Eurygaster integriceps, злаковыми тлями Sitobion avenae F. и Schizaphis graminum Roud., пшеничным трипсом Haplothrips tritici Kurd, пьявицей красногрудой Oulema melanopus L. и хлебными пилильщиками Cephus pygmaeus L. и Trachelus tabidus F., и их энтомофагами – представителями семейств Aelothripidae, Chrysopidae, Coccinellidae, Scelionidae, Syrphidae, Aphidiidae, Encyrtidae, Icehneumonidae, Tachinidae.
Сопряженное развитие этих видов фитофагов и их энтомофагов с озимой пшеницей в весенне-летний период вегетации начинается с фазы выхода в трубку до полной спелости, что охватывает VI–XII этапы органогенеза продуцента, связанные с формированием генеративных, репродуктивных органов и урожая зерна. Это определяет высокую степень вредоносности установленного комплекса вредителей в фазы колошения – восковой спелости, так как наносимые ими повреждения приводят к прямым потерям урожая и снижению его качества, что требует обязательной защиты озимой пшеницы от комплекса вредителей в этот период вегетации.
Биоинсектицид Биослип БВ с нормой расхода 3 л/га, наиболее эффективно подавляет в посевах агроценоза озимой пшеницы злаковых тлей, его средняя биологическая эффективность в отношении данного вредителя составила 78,3 %, а после третьей обработки через 7 дней она составляла 87 %, так же данный препарат обладает средней биологической эффективностью в отношении вредной черепашки и пшеничного трипса, 60,3-68,7 %. Биоинсектицид Биослип БТ, хорошо подходит для защиты посевов от пьявицы красногрудой, так ка его средняя биологическая эффективность составила 84,2 %, что сопоставима с химической защитой, в отношении других видов вредителей его эффективность низкая и колебалась в пределах 27,2-44,2 %. Применение баковой смеси в половинных нормах расхода данных препаратов не целесообразно, так как эффективность данной смеси колебалась в пределах 23,3-40,7 %, что не достаточно для подавления роста популяции фитофагов в посевах озимой пшеницы и ограничения их вредоносности.
Вредоносность фитофагов в посевах озимой пшеницы велика, так как урожайность на контроле ниже на 0,556 т/га по сравнению с химической обработкой в фазу цветения. Количество урожая напрямую зависит от эффективности применяемого биоинсектицида в борьбе с вредными организмами, так наиболее эффективным в период обработок был Биослип БВ, против клопа вредной черепашки, злаковых тлей, пшеничного трипса и хлебных пилильщиков, только в отношении пьявицы красногрудой его эффективность была низкой, в данном варианте наименьшая разница с хозяйственным эталоном 0,09 т/га. В других вариантах с трехкратным применением препарата Биослип БТ и смеси с половинными его с препаратом Биослип БВ снижение в урожайности составило 0,32-0,42 т/га по отношению к эталону.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
- Абдулхаирова С. С. Злаковые тли в условиях центрального и юго-западного Таджикистана : автореф. дис. … канд. биол. наук. Харьков, 1981. 16 с.
- Абдулхаирова С. С. Энтомофаги злаковых тлей // Тезисы докл. участников конференции молодых ученых и профессорско-преподавательского состава Таджикистана. Душанбе, 1977. С. 31-32.
- Агротехнический метод защиты растений / В. А. Чулкина, Е. Ю. Торопова, Ю. И. Чулкин, Г. Я. Стецов. М. : Изд-во ЮКЭА, 2000. 335 с.
- Алексеев A. B. Агроэкологические аспекты защиты озимой пшеницы от пшеничного трипса и злаковых тлей в зоне неустойчивого увлажнения Ставропольского края : автореф. дис. … канд. с.-х. наук. Ставрополь, 2003. 23 с.
- Алексеев A. В., Демкин В. И. Пути снижения вредоносности пшеничного трипса и злаковых тлей // Защита и карантин растений. 2005. № 6. С. 18-20.
- Алехин В. Т. Вредная черепашка // Защита и карантин растений. 2002. № 4. С. 65-91.
- Алехин В. Т. Вредная черепашка и проблема получения качественного зерна // Защита и карантин растений. 2009. № 5. С. 6-7.
- Алехин В. Т. Пути оптимизации защиты зерновых культур // Защита и карантин растений. 2014. № 8. С. 3-8.
- Антропогенная трансформация агроэкосистем и ее фитосанитарные последствия / В. А. Павлюшин, С. Р. Фасулати, Н. А. Вилкова, Г. И. Сухорученко, Л. И. Нефедова. СПб. : ВИЗР, 2008. 120 с.
- Ареал и зоны вредоносности вредной черепашки Eurygaster integriceps Put. (Heteroptera, Scutelleridae) / В. В. Нейморовец, И. Я. Гричанов, Е. И. Овсянникова, М. И. Саулич // Вестник защиты растений. 2006. № 4. С. 2731.
- Артохин К. С. Вредители сельскохозяйственных культур : справочное и учебно-методическое пособие / под общ. ред. К. С. Артохина. М. : ПЕЧАТНЫЙ ГОРОД, 2012. Т. I : Вредители зерновых культур. 532 с.
- Арутюнова Е. В. Энтомофаги основных вредителей озимой пшеницы и кукурузы в северной Осетии // Экономически безопасные и пестицидные технологии получения растениеводческой продукции. Пущино, 1994. С. 90105.
- Баздырев Г. И., Третьяков Н. Н., Белошапкина О. О. Интегрированная защита растений от вредных организмов : учеб. пособие. М. : Инфра-М, 2014. 302 с.
- Баршадская С.И. Урожайность и качество зерна различных сортов озимой пшеницы в зависимости от предшественника, удобрений и других приемов выращивания / С.И. Баршадская, Н.Н. Нещадим, А.А. Квашин // Научный журнал КубГАУ — Scientific Journal of KubSAU. 2016. №120. С.1305-1321
- Белюченко И.С., Боташева Н.Н. Состав основных консортов фито — и энтомофагов озимой пшеницы в засушливой зоне Ставрополья // Экологические проблемы Кубани. 2003. № 19. — С. 37 — 52.
- Белицкая М. Н., Иванцова Е. А. Фауна энтомофагов в лесоаграрных ландшафтах аридной зоны // Вестник Волгоградского государственного университета. Серия 11 : Естественные науки. 2012. № 2. С. 50-55.
- Белошапкин С. П., Шаронов Д. А. Регуляция численности злаковых тлей в посевах озимой пшеницы с помощью агротехнических приемов // Известия Тимирязевской с.-х. академии. 1991. № 6. С. 107-117.
- Белюченко И. С., Боташева Н. Н. Состав основных консортов фито- и энтомофагов озимой пшеницы, в засушливой зоне Ставрополья // Экологические проблемы Кубани : материалы II науч. конф. / НИИ экологии КубГАУ. Краснодар, 2003. С. 37-51.
- Бондаренко Н. В. Биологическая защита растений. М. : Агропромиздат, 1986. 276 с.
- Боташева Н. Н. Влияние условий среды на структуру основных консортов озимой пшеницы и аспекты экологического мониторинга : сб. науч. тр. М., 2002. Т. 1 : Международный форум «Наука III тысячелетия». С. 110-114.
- Боташева Н. Н. Экологическая устойчивость агробиоценоза пшеничного поля в центральной зоне Предкавказья // Физиология человека и животных экологическая безопасность : материалы I Междунар. науч. интернет-конф. / СГСХА. Ставрополь, 2002 а. С. 26-29.
- Бровкин В. И., Соколенко С. Ф. Как повысить урожай озимой пшеницы // Защита и карантин растений. 2010. № 11. С. 20-22.
- Бурлака Г. А., Каплин В.Г. Биологическое обоснование защиты зерновых злаков от хлебных клопов (надсемейства Реи1а1ошо1ёае) в лесостепепи Среднего Поволжья : монография. Кинель : РИЦ СГСХА, 2015. 145 с.
- Васильев В. П. О концепции экономических порогов вредности // Защита растений. 1988. № 1. С. 28-31.
- Васильева Н.Н. Агроэкологические основы защиты озимой пшеницы от стеблевых хлебных пилильщиков в зоне неустойчивого увлажнения : дис. … канд. с-х. наук. Краснодар, 2005. 162 с.
- Викторов Г. А. Проблемы биологической борьбы с вредителями // Защита растений. 1971. № 2. С. 4-6.
- Викторов Г. А. Экология паразитов-энтомофагов. М. : Наука, 1976. 152 с.
- Вилкова Н. А. Воздействие пищеварительных ферментов вредной черепашки на зерновку пшеницы в связи с устойчивостью растений // Труды 5 Всес. Совещ. По иммунитету к болезням и вредителям растений. Киев. 1969 С. 65.
- Вилкова Н. А. Иммунитет растений к вредным организмам и его биоценотическое значение в стабилизации агроэкосистем и повышении устойчивости растениеводства // Вестник защиты растений. 2000, 2, С. 2-15.
- Вилкова Н. А., Ивашенко Л. С. Иммунитет растений к вредителям и его роль в биорегуляции агроэкосистем // Труды русского энтомологического общества. 2001. Вып. 72. С. 129-144.
- Вилкова Н. А., Конарев А. В. Современные проблемы иммунитета растений к вредителям // Вестник защиты растений. 2010. № 3. С. 3-15.
- Вилкова Н.А., Нефедова Л, Капусткина А.В. Характер прорастания зерновок озимой пшеницы, поврежденных вредной черепашкой /II Всерос. совещ. «Современные проблемы иммунитета растений к вредным организмам». СПб: РАСХН, ВИЗР, ИЦЗР, 2008, с. 200-204.
- Воронин К.Е. Использование в биометоде природных популяций энтомофагов // Защита растений. 1977. — № 9. — С. 20-21.
- Воронин К. Е. Охрана окружающей среды при интенсивных технологиях // Защита растений. 1988. № 1 С. 8-10.
- Воронин К. Е., Воронина Э. Г. Факторы, определяющие эффективность энтомофагов и энтомопатогенов // Защита растений. 1996. № 6. С. 14-15.
- Воронин К. Е., Пукинская Е. А., Гусев Е. В. Временные методические указания по использованию критериев эффективности природных энтомофагов. М., 1986. 65 с.
- Воронин К. Е., Шапиро В. А., Пукинская Г. А. Биологическая защита зерновых культур от вредителей. М. : Агропромиздат, 1988. 197 с.
- Вредная черепашка: распространение, вредоносность, методы контроля / В. А. Павлюшин, Н. А. Вилкова, Г. И. Сухорученко, Л. И.Нефедова // Защита и карантин растений. 2010. Т. 1, № 2. С. 54.
- Вредная черепашка и другие хлебные клопы / В.А. Павлюшин, Н.А. Вилкова, Г.И. Сухорученко, Л.И. Нефедова, А.В. Капусткина. СПб., 2015. 280 с.
- Вронских М. Д. Технологии возделывания полевых культур и развитие вредителей и болезней. Кишинев : Pontos, 2005. 290 c.
- Гладков Е. А. Общая экология. М. : МГУИЭ, 2007. 158 с.
- Глазунова Н. Н. Видовое разнообразие и численность энтомофагов на зерновых культурах Ставропольского края // Защита и карантин растений : сб. науч. тр. / СГСХА. Ставрополь, 1998. С. 45-47.
- Глазунова Н. Н. Видовой состав энтомофагов вредной черепашки в центральной зоне Ставропольского края // Проблемы развития биологии и химии на Северном Кавказе : материалы науч.-метод. конф. «Университетская наука — региону» / СГУ. Ставрополь, 1999. С. 48-49.
- Глазунова Н. Н. Динамика численности энтомофагов основных вредителей озимой пшеницы на производственных посевах учебно-опытного хозяйства СГСХА // Химизация растениеводства и вопросы экологии : сб. науч. тр. / СГСХА. Ставрополь, 1999а. С. 57-62.
- Глазунова Н. Н. Пути сохранения и увеличения полезной энтомофауны в агроценозах // Перспективы внедрения современных биотехнологических разработок для повышения эффективности сельскохозяйственною производства : материалы регион. конф. Краснодар, 2000 а. С. 17-20.
- Глазунова Н. Н. Роль экологических факторов в динамике численности основных вредителей и их энтомофагов на озимой пшенице // Защита растений : сб. науч. тр. / СГСХА. Ставрополь, 2000. С. 65-72.
- Глазунова Н. Н. Состав и структура основных консортов озимой пшеницы в зоне неустойчивого увлажнения Ставрополья : монография. Ставрополь : Изд-во СтГАУ «АГРУС», 2004. 104 с.
- Глазунова Н.Н. Уровни сопряженного развития консортов озимой пшеницы // Проблемы экологии и защиты растений в сельском хозяйстве : материалы 68-й науч.-практ. конф. / СтГАУ. — Ставрополь, 2004а. — С. 23 — 29.
- Глазунова Н. Н., Лопатин А. Экологические аспекты поддержания иммунетета агроценоза озимой пшеницы к комплексу факторов // Проблемы экологии и защиты растений в сельском хозяйстве : материалы 68 науч. — практ. конф. / СтГАУ. Ставрополь, 2004б. С. 19-23.
- Глазунова Н. Н. Экологические аспекты биорегуляции сообществ в агроценозе озимой пшеницы // Интегрированная защита сельскохозяйственных культур и фитосанитарный мониторинг в современном земледелии : материалы Всерос. науч.-практ. конф., посвящ. 40-летию факультета защиты растений / СтГАУ. Ставрополь, 2004 в. С. 445452.
- Глазунова Н.Н. Экологическая и экономическая целесообразность применения мобильных критериев ЭПВ для основных вредителей озимой пшеницы // Фитосанитарное оздоровление экосистем. Второй Всерос. съезд по защите растений. (Санкт-Петербург, 5-10 декабря 2005 г.). СПб., 2005. Т. 2. С. 221 — 222.
- Глазунова H. H. Динамика численности злаковых тлей и их энтомофагов в чистосортных и смешанных посевах озимой пшеницы // Проблемы энтомологии Северо-Кавказкого региона : материалы I Региональной науч.- практ. конф. Ставрополь, 2005. С. 92-97.
- Глазунова H. H. Пшеничный трипс и его энтомофаги // Проблемы энтомологии Северо-Кавказкого региона : материалы I Всерос. науч.-практ. интернет-конф. Ставрополь, 2006. Вып. 2. С. 65-67.
- Глазунова Н.Н. Взаимоотношения между видами в консорции озимой пшеницы // Известия высших учебных заведений. Северо-Кавказский регион. Серия: Естественные науки. 2006. № 5. С. 65-70.
- Глазунова Н. Н., Безгина Ю. А., Устимов Д. В. Математическое описание взаимосвязи численности вредной черепашки, теленомин, фазий и погодно- климатических факторов фазы онтогенеза озимой пшеницы // Вестник АПК Ставрополья. 2013. № 4 (12). С. 160-169.
- Глазунова Н. Н., Безгина Ю. А., Устимов Д. В. Математическое моделирование, изменения численности популяции злаковых тлей, и её энтомофагов (паразитов и хищников) в разные периоды онтогенеза озимой пшеницы и погодно-климатических факторов // Современные проблемы науки и образования. 2013 а. № 6. С. 965.
- Глазунова Н. Н. Математическое моделирование взаимосвязи погодно- климатических факторов и динамики численности популяции Cephus pygmaeus L. и Collyria coxator Vill. в разные фазы онтогенеза озимой пшеницы в Ставропольском крае // Современные проблемы науки и образования. 2014 а. № 2. С. 601.
- Глазунова Н. Н. Новый подход к защите от основных вредителей в весенне-летний период // Труды Ставропольского отделения Русского энтомологического общества / СтГАУ. Ставрополь, 2014. Вып. 10 : материалы VII Междунар. науч.-практ. интернет-конф. «Актуальные вопросы энтомологии». С. 66-75.
- Глазунова Н.Н., Шутко А.П., Безгина Ю.А., Тутуржанс Л.В. и др. Методические рекомендации «Система защиты озимой пшеницы от вредителей и болезней на Юге России». Ставрополь: СЕКВОЯ, 2018. 98с.
- Действие и взаимодействие минеральных удобрений и пестицидов и влияние их на фитосанитарное состояние посевов озимой пшеницы / Н. А. Цветкова, Р. А. Федорова, В. Ф. Ладонин, А. М. Алиев, А. М. Симон // Сб. науч. тр. ВИЗР. СПб., 1995. № 6. С. 18-37.
- Демкин В. И., Глазунова Н. Н. Агроэкологические аспекты защиты урожая зерновых и зернобобовых в условиях Центрального Предкавказья // Сб. науч. тр. межрегион., межвуз. конф. / СтГАУ. Ставрополь, 2000. С. 100-101.
- Добронравова М. В., Глазунова Н. Н. Фитосанитарное состояние и защита озимой пшеницы от сосущих вредителей в Центральном Предкавказье // Современные проблемы науки и образования. 2013. № 3. С. 425.
- Дорожко Г. Р., Власова О. И., Передериева В. М. Способ обработки — фактор регулирования фитосанитарного состояния почвы и посевов озимой пшеницы на черноземах выщелоченных зоны умеренного увлажнения Ставропольского края // Научный журнал Кубанского государственного аграрного университета : политематический сетевой электронный журнал. 2011. № 68. С. 442-450.
- Дорожко Г.Р., Власова О.И., Цховребов В.С. Развитие земледелия Ставрополья // Эволюция и деградация почвенного покрова : материалы V Междунар. науч.-практ. конф. Ставрополь, 2017. С. 249-251.
- Доспехов Б. А. Методика полевого опыта (с основами статистической обработки результатов исследований). М. : Агропромиздат, 1985. 351 с.
- Журавлёва Е.Н. Влияние предшественников и основной обработки почвы на агроценоз озимой пшеницы в умеренно влажной зоне: дис. … канд. с-х. наук. Ставрополь, 2002. 153 с.
- Жученко А. А. Адаптивное растениеводство (эколого-генетические основы). Кишинев : Штиница, 1990. 432 с.
- Жученко А. А. Стратегия адаптивной интенсификации сельского хозяйства (концепция). Пущино : ОНТИ ПНЦ РАН, 1994. 148 с.
- Жученко А. А. Экологическая генетика культурных растений как самостоятельная научная дисциплина (теория и практика). Краснодар: Просвищение-Юг, 2000. 485 с.
- Жученко А. А. Экологическая генетика культурных растений и проблемы агросферы (теория и практика). М.: Изд. Агрорус., 2004. Т. 1- 690 с., 2-466 с.
- Жученко А.А. Экологическая генетика культурных растений как самостоятельная научная дисциплина. Теория и практика. — Краснодар: Просвещение Юг, 2010, 485 с.
- Зазимко М. И. Концептуальные основы экологизации системы защиты колосовых культур на Северном Кавказе // Экологически безопасные и беспестицидные технологии получения растениеводческой продукции : материалы Всерос. науч.-производ. совещ. Пущино, 1994. С. 17-19.
- Зазимко М. И. Экологизированная система защиты колосовых культур от болезней и вредителей на Северном Кавказе : дис. … д-ра с.-х. наук. СПб., 1995. 415 с.
- Зазимко М. И., Гузь А. Л., Лактионова Н. В. Основы экологизации защиты колосовых культур в Северо-Кавказком регионе : тез. докл. Всерос. съезда по защите растений. СПб., 1995. С. 209-214.
- Замотайлов А.С. Жужелицы (Coleoptera, Carabidae) в агроценозах Краснодарского края // Труды Кубанского ГАУ «Защита растений от болезней, вредителей и пути повышения устойчивости к ним», 1993, 332 (360), С. 4-13.
- Захаренко В. А. Фитосанитарные риски в зерновом производстве // Защита и карантин растений. 2014. № 7. С. 3-7.
- Захаренко В. А., Сударенков Г. В. Экологическая оценка фитосанитарного состояния агроэкосистем // Фитосанитарная ситуация на посевах сельскохозяйственных культур юга России и экологизация систем защиты : материалы науч.-практ. конф. Краснодар, 2000. С. 8-9.
- Защита зерновых культур на основе фитосанитарной оценки / Ю. Б. Шуровенков, Ю. В. Попов, О. В. Попова, В. Ф. Рукин // Защита и карантин растений. 1998. № 10. С.13-15.
- Интегрированные системы зашиты сельскохозяйственных культур от вредителей, болезней и сорняков. Рекомендации. В 2-х книгах / под ред. С. В. Сорока. Минск : УП «ИВС Минфина». 2003. Кн. 1. 249 с.
- Интегрированные системы зашиты сельскохозяйственных культур от вредителей, болезней и сорняков. Рекомендации. В 2-х книгах / под ред. С. В. Сорока. Минск : УП «ИВС Минфина». 2003. Кн. 2. 256 с.
- Карпенко Я. Г. Охрана полезных членистоногих // Защита растений. 1996. № 2. С. 11-12.
- Коваленков В. Г. Состояние проблемы резистентности к пестицидам вредных организмов и пути перехода к биоценотическому контролю её развития в условиях Еверо-Кавказского региона / Материалы конференции / Российская академия сельскохозяйственных наук, Всероссийский научно- исследовательский институт биологической защиты растений. 2000. С. 1221.
- Коваленков В. Г., Тюрина Н. М. Как умножить и сохранить полезную биоту // Защита растений. 2016. № 10. С. 25-26.
- Математическое описание взаимосвязи динамики численности популяции Haplothrips tritici Kurd. и Aelothrips fasciatus L. в разные фазы онтогенеза озимой пшеницы от погодно-климатических факторов в Центральном Предкавказье / Н. Н. Глазунова, Ю. А. Безгина, Л. В. Мазницына, О. В. Шарипова, Д. В. Устимов // Современные проблемы науки и образования. 2014. № 1. С. 379.
- Мейер Н. Ф. Биологический метод борьбы с вредными насекомыми. М.- Л. : Сельхозгиз, 1937. 188 с.
- Методика государственного сортоиспытания сельскохозяйственных культур. Вып. 2 : Зерновые, зернобобовые, кукуруза и кормовые культуры. М. : Колос, 1989. 294 с.
- Методика количественной оценки роли энтомофагов в полевых условиях (унифицированный подход) / сост. А. Ф. Зубков ; Л. : ВИЗР, 1984. 20 с.
- Методы определения болезней и вредителей сельскохозяйственных растений / И. Беттхер, Т. Ветцель, Ф. В. Древс [и др.] ; пер. с нем. К. В. Попковой, В. А. Шмыли. М. : Агропропиздат, 1987. 223 с.
- Методы оценки сельскохозяйственных культур на групповую устойчивость к вредителям / Н. А. Вилкова, Б. П. Асякин, Л. И. Нефедова [и др.]. СПб. : ВИЗР, 2003. 112 с.
- Мизер А. В., Посылаева Г. А., Янова Т. А. Пути сохранения энтомофагов злаковых тлей // Исследования по энтомологии и акарологии на Украине : тез. докл. II съезда УЭО. Киев, 1980. С. 194-195.
- Маланина З. И., Колмукиди С. В., Крюкова Е. А. Роль сорной травянистой и древесно-кустарниковой растительности в инфекционных процессах аграрных и агролесных ландшафтов // Защита и карантин растений. 2011. №4.
- Мохрин А. А., Ченикалова Е. В. Мониторинг и прогноз численности и эффективности кокцинеллид в борьбе с вредителями в агробиоценозах Ставропольского края // Интегрированная защита сельскохозяйственных культур и фитосанитарный мониторинг в современном земледелии : сб. науч. тр. по матер. Междунар. науч.-практ. конф. / СтГАУ. Ставрополь, 2007. С. 246-249.
- Мухина О. В., Шабалдас О. Г., Солодилина А. Ю. Влияние морфофизиологических особенностей сортов озимой пшеницы на устойчивость к важнейшим вредителям // Применение современных ресурсосберегающих инновационных технологий в АПК : сб. тр. по материалам Междунар. науч.-практ. конф. Ставрополь, 2013. С. 148-152.
- Надыкта В. Д., Исмаилов В. Я., Коваленков В. Г. Биологическая борьба с вредителями: ученые предлагают. Дело за внедрением // Защита и карантин растений. 2007. № 4. С. 19-21.
- Новожилов К. В. Фитосанитарная оптимизация растениеводства // Защита и карантин растений. 1996. № 8. С. 15-17.
- Новожилов К. В. Проблемы оптимизации фитосанитарного состояния растениеводства // Сельскохозяйственная биология. 1997. № 5. С. 28-38.
- Новожилов К. В. Некоторые направления экологизации защиты растений // Защита и карантин растений. 2003. № 8. С. 14-17.
- Новожилов К. В. Аспекты повышения экологичности фитосанитарных блоков в технологиях современного растениеводства / Роль и место с.-х. науки в агропромышленном комплексе России. Юбилейная сессия РАСХН: Генитические ресурсы и биотехнология, М.,2005. С. 84-92.
- Новожилов К.В., Захаренко В.А. Принципы, критерии и технологии стабилизации фитосанитарного состояния агроэкосистем. СПб., 2000 . 99с.
- Новожилов К.В., Захаренко В.А. Подходы к конструированию агроэкосистем — интегрирование методов и средств защиты растений с целью управления фитосанитарным состоянием сельскохозяйственных культур. СПб. : ВИЗР, 2000 а. 92 с.
- Новожилов К. В., Захаренко В. А. Уровни и тенденции изменения видового состава и внутренней структуры, ареалы комплексов вредных и полезных организмов и прогноз опасных фитосанитарных ситуаций по зонам страны. СПб. : ВИЗР, 2000 б. 100 с.
- Новожилов К.В., Сухорученко Г.И. Экологические принципы использования инсектоакарицидов в сельском хозяйстве // Агрохимия. 1995. № 1. С. 111-118.
- Новожилов К. В., Сухорученко Г.И. Методические принципы оценки степени опасности инсектоакарицидов для полезных членистоногих / Сборник трудов Всероссийского съезда по защите растений «Проблемы оптимизации фитосанитарного состояния растениеводства». СПб., 1997. С. 281-286.
- Осмоловский Г.Е. Выявление сельскохозяйственных вредителей и сигнализация сроков борьбы с ними. М.: Россельхозиздат, 1964. 204 с.
- Оценка экологической опасности пестицидов для агробиоценозов / К. В. Новожилов, Г.И. Сухорученко, Н.Н. Семенова С.А Волгаре, В.М Питулько // Региональная экология РАН. 2010 а. № 1-2 (28). С. 73-79.
- Павлов И.Ф. Агротехнические и биологические методы защиты растений. М. : Россельхозиздат, 1976. 280 с.
- Павлюшин В.А. Фитосанитарные последствия антропогенной трансформации агроэкосистем / В.А. Павлюшин, Н.А. Вилкова, Г.И. Сухорученко, С.Р. Фасулати, Л.И. Нефедова // Вестник защиты растений. 2008. № 3. С. 3-26.
- Павлюшин В.А. Резистентность вредных членистоногих к пестицидам и меры её преодоления / В.А. Павлюшин, К.В. Новожилов, Н.А. Вилкова, Г.И. Сухорученко // Защита и карантин растений. 2013. № 5. С. 62-89.
- Павлюшин В.А. Фитосанитарная оптимизация агроэкосистем / В.А. Павлюшин, К.В. Новожилов, Н.А. Вилкова, Г.И. Сухорученко // В сборнике: Фитосанитарная оптимизация агроэкосистем Материалы III Всероссийского съезда по защите растений. 2013 а. С. 150-158.
- Павлюшин В.А. Интегрированная защита озимой пшеницы / В.А. Павлюшин, В.И. Долженко, А.М. Шпанев, А.Б. Лаптиев, Н.Р. Гончаров, А.К. Лысов, О.В. Кунгурцева, Л.Д. Гришечкина, Л.А. Буркова, А.С. Голубев, А.А. Яковлев, Н.В. Бабич, А.И. Силаев, В.А. Хилевский, Н.Н. Лунева, Т.Ю. Гагкаева, Н.А. Вилкова, Л.И. Нефедова, Г.И. Сухорученко, Е.И. Гультяева и др. // Защита и карантин растений. 2015. № 5. С. 38-71.
- Павлюшин В.А. Роль иммунитета растений в фитосанитарной оптимизации агроэкосистем (Новая парадигма защиты растений) / В.А. Павлюшин, Н.А. Вилкова, Г.И. Сухорученко, Л.И. Нефедова // Современные проблемы иммунитета растений к вредным организмам : Тезисы докл. IV Междунар. науч. конф. 2016. С. 4-6.
- Павлюшин В.А. Современные проблемы управления фитосанитарным состоянием агроэкосистем / В.А. Павлюшин, Н.А. Вилкова, Г.И. Сухорученко, Л.И. Нефедова // Информационный бюллетень ВПРС МОББ. 2017. № 52. С. 221-227.
- Павлюшин В.А. Фундаментальные исследования в области сельскохозяйственной энтомологии в решении проблем фитосанитарной оптимизации агроэкосистем Современные проблемы управления фитосанитарным состоянием агроэкосистем / В.А. Павлюшин, Н.А. Вилкова, Г.И. Сухорученко, Л.И. Нефедова // В сборнике: XV Съезд Русского энтомологического общества Материалы съезда. 2017а. С. 379-380.
- Пикушова Э. А., Веретельник Е. Ю., Пшидаток С. К. Влияние способов обработки почвы на численность основных вредителей озимой пшеницы сорта Фортуна // Агротехнический метод защиты растений от вредных организмов : материалы V науч.-практ. конф. / КубГАУ. Краснодар, 2011. С. 190-195.
- Поляков И. Я. Прогноз развития вредителей и болезней сельскохозяйственных культур (с практикумом) / И. Я. Поляков, М. П. Персов, В. А. Смирнов Л. : Колос, 1984. 318 с.
- Поляков И.Я. Методические рекомендации по разработке прогнозов динамики популяций вредителей и планированию объемов защитных мероприятий на основе агроклиматических предикторов. /И.Я. Поляков Л.А. Макарова, Г.М. Доронина. — Л.: ВИЗР, 1985, 51 с.
- Прогноз фитосанитарного состояния посевов сельскохозяйственных культур Ставропольского края на 2017 годи системы защитных мероприятий : рекомендации для сельхозтоваропроизводителей / под общ. ред. П.Д. Стамо. Ставрополь. : Бюро новостей, 2017. 184 с.
- Прогноз фитосанитарного состояния посевов сельскохозяйственных культур Ставропольского края на 2015 годи системы защитных мероприятий : рекомендации для сельхозтоваропроизводителей / под общ. ред. П.Д. Стамо. Ставрополь. : Бюро новостей, 2015. 164 с.
- Пукинская Г. А. Использование природных популяций энтомофагов в интегрированной борьбе с тлями на зерновых культурах // Интегрированная защита зерновых культур. М., 1981. С. 112-119.
- Пукинская Г. А. Эффективность энтомофагов злаковых тлей // 8-й Междунар. конгресс по защите растений : тез. докл. советских участников конгресса. М., 1975. С. 178-179.
- Самерсов В. Ф. Интегрированная система защиты зерновых культур от вредителей. М. : Урожай, 1988. 204 с.
- Системы интегрированной защиты посевов озимой пшеницы от вредных организмов в Северо-Западном регионе РФ / А.М. Шпанев, А.Б. Лаптиев, Н.Р. Гончаров, А.С. Голубев, Т.А. Маханькова, О.В. Кунгурцева, Е.И. Гультяева, Н.П. Шипилова, Н.Н. Лунева, Е.Н. Мысник, О.А. Баранова, Н.М. Коваленко, О.Г. Гусева, П.В. Лекомцев, В.В. Воропаев, В.В. Смук ; под общ. ред. В.А. Павлюшина. — СПб., ВИЗР, 2017. — 47 с.
- Системы земледелия Ставрополья [Электронный ресурс] : моногр. / А. А. Жученко, В. И. Трухачев, В. М. Пенчуков, В. С. Цховребов, В. М. Передериева, О. И. Власова, А. Н. Есаулко, В. В. Агеев, А. И. Подколзин, О. Ю. Лобанкова, Г. Р. Дорожко, О. Г. Шабалдас, Т. Г. Зеленская, В. С. Сотченко, В. Н. Багринцева, В. К. Дридигер, Г. П. Полоус, В. Г. Гребенников, М. П. Жукова, А. И. Войсковой, Н. З. Злыднев, Р. М. Злыднева, О. Г. Ангилеев, А. Ю. Раков, А. А. Сентябрев, М. А. Сирота ; под общ. ред. А. А. Жученко, В. И. Трухачева. Ставрополь : Изд-во СтГАУ «АГРУС», 2011.
- Скребцова Т. И. Биоэкологические особенности вредной черепашки (Eurygaster integriceps PUT.) и совершенствование мер борьбы с ней в Центральном Предкавказье : автореф. дисс. … канд. с-х. наук. М., 2009. 25 с.
- Скребцова Т. И. Ресурсосберегающие технологии и защита озимых культур // Защита и карантин растений. № 5. 2008. С. 7-9.
- Скребцова Т. И., Ченикалова Е. В., Пентык И. Д. Динамика распространения и численности вредной черепашки в Ставропольском крае // Защита и карантин растений. 2009. № 9. С. 44.
- Современные экологически безопасные системы фитосанитарной оптимизации растениеводства в Сибири (теория, методология, практика) / В. А. Чулкина, Е. Ю. Торопова, В. М. Медведчиков, Г. Я. Стецов, Ю. И. Чулкин [и др.]. Новосибирск, 2003. 116 с.
- Соколов М. С. Экологизированное растениеводство как фактор устойчивого развития АПК России // Вестник защиты растений. 2001. Т. 1. С. 63-70.
- Соколов М. С., Монастырский О. А., Пикушова Э. А. Экологизация защиты растений. Пущино : ОНТИ ПНЦ РАН, 1994. 462 с.
- Соколов М. С., Торонова Е. Ю., Чулкина В. А. Общие принципы разработки и реализации фитосанитарных технологий // Вестник защиты растений. 2007. № 2. С. 25-43.
- Сухорученко Г.И., Новожилов К.В. Приоритеты токсикологических исследований на современном этапе химического метода защиты растений / Химический метод защиты растений: Состояние и перспективы повышения экологической безопасности. СПб., 2004. С. 308-312.
- Сухорученко Г.И., Долженко В.И., Гончаров Н.Р., Васильева Т.И., Иванов С.Г., Иванова Г.П., Буркова Л.А., Тайманов Ш.И., Зенькевич С.В., Зверев А.А., Белых Е.Б. Технология и методы оценки побочных эффектов от пестицидов (на примере преодоления резистентности колорадского жука к инсектицидам). СПб., 2006. 52 с.
- Танский В. И. Биоценотический подход к интегрированной защите растений от вредных насекомых // Энтомологическое обозрение. 1997. № 2. С. 251-264.
- Танский В. И. Экономические пороги вредоносности насекомых и их роль в защите растений : информ. бюл. В.П.С. МОББ. СПб., 1981. 93 с.
- Танский В. И., Тулеева А. К. Влияние предшественников на вредных и полезных насекомых в агроценозах яровой пшеницы // Вестник защиты растений. 2005. № 1. С. 27-31.
- Теленга Н. А. О роли энтомофагов в массовых размножениях насекомых // Зоологический журнал. 1953. Т. 32, вып. 1. С. 14-24.
- Тильменбаев А.Т. Роль яйцеедов в регулировании численности вредной черепашки в Казахстане / Краткие тезисы докладов к совещанию по проблемам биологической борьбы с вредной черепашкой в интегрированной системе защиты зерновых культур. Л.: ВИЗР, 1971. С. 114-116.
- Титова Ю. А. Биоценотическая регуляция — основа современных фитосанитарных технологий // Защита и карантин растений. 2007. № 12. С. 44-45.
- Тряпицын В. А., Шапиро В. А., Щепетильникова В. А. Паразиты и хищники вредителей сельскохозяйственных культур. Л. : Колос, 1982. 256 с.
- Устойчивость зерновых культур к вредителям и их вредоносность на современных сортах озимой пшеницы / Е. В. Ченикалова, О. В. Мухина, С.А. Щербакова, Т. И. Скребцова, Ю. А. Безгина., С. И. Любая. Ставрополь, 2008. 108 с.
- Фитосанитарная дестабилизация агроэкосистем / В.А. Павлюшин, Н.А. Вилкова, Г.И. Сухорученко, Л.И. Нефедова, С.Р. Фасулати. СПб. : НППЛ «Родные просторы», 2013. 184 с.
- Фокин А. В. Эффект матрешки в теории биометода // Защита и карантин растений. 2014. № 4. С. 52-53.
- Химическая защита растений в фитосанитарном оздоровлении агроэкосистем / В. И. Долженко, К. В. Новожилов, Г. И. Сухорученко, С. Л. Тютерев // Вестник защиты растений. 2011. № 3. С. 3-12.
- Холодковский Н. А. Второй каталог коллекции тлей (Aphididae) // Известия Санкт-Петербургского лесного института. 1902. Вып. 8. С. 49-61.
- Холодковский Н. А. Жизнь тлей и значение их в биологии // Природа. 1914. № 12. С. 1423-1440.
- Холодковский Н. А. Второй каталог коллекции тлей (Aphididae) // Известия Санкт-Петербургского лесного института. 1902. Вып. 8. С. 49-61.
- Чекмарева Л.И. Изменение элементов агроценоза пшеницы под влиянием обработки почвы /Л.И. Чекмарева, Е.П. Денисов, С.Г. Лихацкая, И.С. Полетаев, Д.М. Лихацкий // «Известия Оренбургского государственного аграрного университета». — № 3 (53). — 2015. — С. 396
- Чернышев В. Б. Сельскохозяйственная энтомология (экологические основы) : курс лекций. М. : Триумф, 2012. 232 с.
- Чертова Т. С. Экологические проблемы защиты растений // Защита и карантин растений. 1997. № 9. С. 44.
- Чулкина В. А., Торопова Е. Ю., Стецов Г. Я. Экологические основы интегрированной защиты растений. М. : Колос, 2007. 568 с.
- Чулкина В. А., Чулкин Ю. И. Управление агроэкосистемами в защите растений. Новосибирск, 1995. 202 с.
- Шадрина Л. А. Эффективность биологических и химических средств защиты озимой пшеницы от вредных организмов на фоне различных приемов возделывания : автореф. дис. … канд. биол. наук. Краснодар, 1996. 24 с.
- Шапиро В. А. Факторы определяющие эффективность энтомофагов в агробиоценозах // Сельскохозяйственная биология. 1991. №3. С. 141-152.
- Шерамеев А., Букарев К., Рузиев А. Влияние предшественников на фитосанитарное состояние и урожай озимой пшеницы // Защита и карантин растений. 2001. № 8. С. 20.
- Ширинян Ж.А., Исмаилов В.Я. Роль природных популяций энтомофагов в ограничении численности вредной черепашки (Erygaster integriceps Put.) (Hemiptera, 8с^е11епёае)на посевах озимой пшеницы в условиях Юга России и оптимизация защитных мероприятий / Сб. научн. трудов. «Биологическая защита растений — основа стабилизации агроэкосистем». Краснодар, 2004. С. 105-112.
- Шпаар Д. Зерновые культуры. Выращивание, уборка, хранение и использование / под общ. ред. Д. Шпаара. М. : Зерно, 2012. 704 с.
- Шутко А.П. Биологическое обоснование оптимизации системы защиты озимой пшеницы от болезней в Ставропольском крае : автореферат дис. … доктора сельскохозяйственных наук : 06.01.07 / Всероссийский научно-исследовательский институт защиты растений Российской академии сельскохозяйственных наук. Санкт-Петербург-Пушкин, 2013
- Шутко А.П., Гаврилов А.А., Передериева В.М. Управление патологическим процессом корневых гнилей на Ставрополье // Вестник АПК Ставрополья. 2011. № 3. С. 16-21.
- Щепетильникова В. А. Закономерности, определяющие эффективность энтомофагов // Журнал общей биологии. 1957. Т. 18, № 5. С. 381-394.
- Щербакова С. А., Мохрин А. А. Влияние сортов озимой пшеницы на заселенность ее злаковой тлей и кокцинеллидами // Молодежная аграрная наука : состояние, проблемы и перспективы развития. Ставрополь, 2007. С. 143-145.
- Яркулов Ф. Я. Как мы используем природных энтомофагов // Защита и карантин растений. 1995. № 2. С. 4-6.
- Яхонтов В. В. Экология насекомых. — М.: Высшая школа, 1969—488с.
- Alford D. V. Pest and Disease Management Handbook // British Crop Protection Council. Blackwell Science. Oxford, 2000. 615 p.
- Aukema В., Rieger Ch. Catalogue of the Heteroptera of Palaearctic Region. Pentato-morpha II // The Netherlands Entomol. Soc. 2005. 550 p.
- Beckage N. E., Gelman D. B., Wasp parasitoid disruption of host development: implications for new biologically based strategies for insect control // Annu. Rev. Entomol. 2004. № 49. P. 299-330.
- Blackwell M. Minute mycological mysteries: the influence of arthropodes on the lives of fungi // Mycologia. 1994. V. 86. № 1. P. 1-17.
- Boivin G., Hance T., Brodeur J. Aphid parasitoids in biological control // Can J Plant Sci. 2012. № 92. P. 1-12.
- Bottcher O., Gent R. Chancen und Risiken von Pflanzenschutzmitteln — der Abwagungsprozess ftlr die Zulassungsentscheidung und Risikokommunation // Mitt Biol Bundesanst. Land- und Forstwirtschaft, heft 371. 2000. P. 36-45.
- Ceolin Bortolotto O, de Oliveira Menezes Jtoior A, Thibes Hoshino A. Aphidophagous Parasitoids can Forage Wheat Crops Before Aphid Infestation, Parana State, Brazil. Journal of Insect Science. 2015;15(1):40.
- Colinet H., Boivin G. Insect parasitoids cold storage: A comprehensive review of factors of variability and consequences // Biol Control. 2011. № 58 P. 83-95.
- Dean G.J. Bionomics of aphids reared on cereal and some Gramineae / G.J. Dean // Ann. appl. Biol. 1973. — Vol. 73, №1. -P. 127-135.
- Dedryver C.-A., Ralec A. Le. Fabre F. The conflicting relationships between aphids and men: a review of aphid damage and control strategies // Comptes Rendus Biologies. 2010. Vol. 333, № 6-7. P. 539-553.
- Grapel D. G., Heinrich J. G. Untersuchungen ur wirtschaftlichen Scha- densschwelle von Getreideblattlausen und zur Wirking einiger Inse rtizide au naturlihe Blactlausfeinde / D; ss…. Gieben. 1980. 134 p.
- Kokubu H., Duelli P. Aerial population movement and vertical distribution of aphido-phagous insects in corn fields (Chrysopidae, Coccinellidae and Syrphidae) // Ecol. Aphidophaga. 1986. V. 2. P. 279-284.
- Kreye H., Garbe V., Bartels G. Pflanzenschutz im Weizen bei konservierender Bodenbearbeitung // Getreide Mag. 1999. Vоl. 5, № 4. P. 180-183.
- Mawela K. V., Kfir R., Krueger K. Effect of temperature and host species on parasitism, development time and sex ratio of the egg parasitoid Trichogrammatoidea lutea Girault (Hymenoptera: Trichogrammatidae) // Biol Control. 2013. № 64. P. 211-216.
- Olanca B., Sivri Ozay D., Koksel H. Effects of suni-bug (Eurygaster spp.) damage on size distribution of durum wheat (Triticum durum L.) proteins // European Food Res. Technol. 2009. № 229. P. 813-820.
- Smith L. Temperature inFLuences functional response of Anisopteromalus calandrae (Hymenoptera: Pteromalidae) parasitizing maize weevil larvae in shelled corn // Ann. Entomol. Soc. Am. 2004. № 87, P. 849-855.
- Smith R.W. Status in Ontario of Collyria calcitrator (Grav.) (Hymenoptera: Ichneumonidae) and of Pediobius beneWcus (Gahan) (Hymenoptera: Eulophidae) as parasites of the European wheat stem sawXy, Cephus pygmaeus (L.) (Hymenoptera: Cephidae) // Can. Entomol. 1959. № 91. P. 697-700.
- Thompson W. R. The time factor in biological control // Canad. Ent. 1951. № 83. P. 230-240.