
Титульный лист и исполнители
РЕФЕРАТ
Отчет 286 с., 105 табл., 36 рис., 251 источников литературы.
МИКРОБИОМ ЖКТ ПТИЦ, БАКТЕРИАЛЬНЫЕ ШТАММЫ, СВОЙСТВА МИКРООРГАНИЗМОВ, ВЫДЕЛЕНИЕ, БИОПРЕПАРАТЫ, БАКТЕРИОЦИНЫ, ПОСЛЕДОВАТЕЛЬНОСТИ, АНАЛИЗ СТРУКТУРЫ, ПОЛИСАХАРИДЫ, ПЕКТИН, АЛЬГИНАТ НАТРИЯ, КСАНТАН, КОРМОВАЯ ПРОБИОТИЧЕСКАЯ ДОБАВКА, ОЦЕНКА КАЧЕСТВА, ТОКСИЧНОСТЬ, ЭФФЕКТИВНОСТЬ ПРИМЕНЕНИЯ, ПЕРЕПЕЛА, ЦЫПЛЯТА-БРОЙЛЕРЫ, ПРОДУКТИВНОСТЬ, СОХРАННОСТЬ, КРОВЬ, КАЧЕСТВО ПРОДУКЦИИ, РЕНТАБЕЛЬНОСТЬ.
В ходе выполнения проекта продемонстрирована важность применения информации, полученной в ходе полногеномного секвенирования пробиотических штаммов бактерий ‒ компонентов сельскохозяйственных биопрепаратов. Данные полногеномного секвенирования, такие, как уточненная информация о таксономической принадлежности штаммов, последовательности генов, отвечающих за проявление пробиотических свойств, кислотообразование, антогонистическую активность по отношению к патогенным и условно-патогенным штаммам (в первую очередь, структура бактериоцинов), необходимы при оптимизации характеристик инновационных биопрепаратов для выращивания сельскохозяйственной птицы.
Получены данные о полных геномах нескольких пробиотических штаммов кисломолочных бактерий, входящих в состав ранее разработанных препаратов «Галлобакт-Ф» и «Галлобакт-ЛТ». Исследования проведены методом высокоэффективного полногеномного секвенирования на секвенаторе нового поколения Illumina MiSeq. Полногеномные последовательности лактобактерий депонированы в международную базу данных NCBI. Уточнена таксономическая принодлежность компонентов препаратов – штаммов Lactobacillus salivarius 16РС20 и Lactobacillus curvatus 13PC723.
Впервые идентифицированы и депонированы в международную базу данных последовательности девяти бактериоцинов. Изучена их структура и выяснена информационная организация отдельных участков молекул.
В работе для оптимизации характеристик новой пробиотической добавки применяли полисахариды различного происхождения, в частности пектин, полученный из продуктов растительного сырья, альгинат натрия, выделенный из водорослей и ксантан, полученный из микробной массы. При подборе оптимальной дозы полисахарида с целью максимальной выживаемости лактобацилл выявлено, что полисахаридная смесь на основе альгината натрия из расчета 0,3 % в составе нового пробиотика, обеспечивает наилучший результат.
Разработан состав и технология получения новой пробиотической добавки «Галлобакт-Ф-Плюс». Отдельно изучены биологические свойства лактобацилл, используемых в составе пробиотика, а именно спектр сбраживаемости сахаров, потребность в отдельных витаминах, активное кислотообразование. Изучен фенотипический профиль антибиотикорезистентности лактобактерий, антагонизм к эталонной условно-патогенной и патогенной микрофлоре, а также к нормофлоре кишечника, антиадгезивные и адгезивные свойства молочнокислых бактерий к клеточным моделям.
Проведено изучение лечебно-профилактического свойства разработанного пробиотика «Галлобакт-Ф-Плюс» на модели экспериментального дисбактериоза у лабораторных животных. Продемонстрировано токсикологическое и раздражающее действие пробиотической добавки на организм лабораторных мышей, крыс и кроликов-альбиносов. Установлено, что доза пробиотика «Галлобакт-Ф-Плюс»: 0,5 мл/гол. – в мясном перепеловодстве и 0,75 мл/гол. – в бройлерном птицеводстве, способствует повышению сохранности птицы, увеличению массы тушки, снижению конверсии комбикорма, улучшению показателей биополноценности продукции птицеводства, а также повышению рентабельности ведения данной отрасли животноводства.
ВЕДЕНИЕ
В кишечнике здоровых птиц содержится около 900 видов микроорганизмов, и их совокупность рассматривается как микробная экологическая система. Существование животных без нормальной микрофлоры желудочно-кишечного тракта (ЖКТ) невозможно, так как она контролирует многие метаболические функции хозяина. Поддерживать нормофлору в стабильном состоянии и регулировать ее состав можно введением в корм пробиотиков – живых организмов, которые при введении в физиологических количествах приносят пользу организму хозяина. Микроорганизмы, используемые в качестве пробиотиков, большинство микробиологов относят к нескольким группам (Lactobacillus, Enterococcus и др.). Традиционно наиболее широко применяются молочнокислые бактерии. В современной научной и технической литературе сформированы четкие требования к потенциальным пробиотическим штаммам. Поскольку одним из основных механизмов, направленных против патогенных бактерий, является конкурентоспособность нормофлоры в борьбе за экологическую нишу, применение представителей автохтонной микрофлоры, входящей в естественные для данного вида микробные ассоциации и, как следствие, обладающие повышенной функциональной адаптацией к физиологическим особенностям выращиваемой птицы, представляется более оправданным.
Учитывая особенности географического расположения промышленных птицеводческих хозяйств, перспективно использование зональных природных резервуаров микроорганизмов, сформировавшихся в конкретных климатических и физико-географических условиях Юга России. Генетическая селекция по признакам высокой продуктивности поголовья сельскохозяйственных птиц
в настоящее время привела к понижению естественной резистентности организма и, как следствие, широкому использованию противомикробных препаратов в составе кормов.
Помимо профилактического и терапевтического действия по отношению
к различным болезням бактериальной этиологии, они также используются как эффективные кормовые добавки, стимулирующие рост молодняка и улучшающие целый ряд зоотехнических параметров (сохранность, продуктивность, коэффициент конверсии корма и др.). Из-за постоянного воздействия антибиотиков патогенные и условно патогенные бактерии в течение времени вырабатывают устойчивость к ним, изменяется состав микрофлоры кишечника (снижается численность полезной кишечной микрофлоры), нарушаются физиологические и иммунологические механизмы защиты организма птиц, создаются условия для развития инфекционных процессов, вызываемых его собственной патогенной микрофлорой и бактериями из окружающей среды. Генетические детерминанты антибиотикорезистентности, попадая в геномы патогенных видов бактерий, а затем и в человека, делают вызываемые ими инфекционные заболевания практически неизлечимыми существующими медикаментами, повышается количество аллергических и аутоиммунных заболеваний у людей.
Одной из рассматриваемых альтернатив широкому применению антибиотиков считаются пробиотические препараты. Одними из наиболее перспективных видов-пробиотиков рода Lactobacillus могут выступать L. salivarius
и L. curvatus. Протокол исследования коммерчески интересных штаммов
в настоящее время предполагает углубленное исследование их генома и транскриптома (как минимум – проведение полногеномного секвенирования).
В целом хотелось отметить, что глубокое изучение научной и патентной литературы показало отсутствие эффективных конструкций функционально-адаптированных пробиотических добавок, изготовленных с применением эволюционно-закрепленных микроорганизмов желудочно-кишечного тракта птицы для применения в промышленном птицеводстве.
1 ОБЗОР ЛИТЕРАТУРЫ
1.1 МИКРОФЛОРА ЖЕЛУДОЧНО-КИШЕЧНОГО ТРАКТА ПТИЦ. МЕТОДЫ АНАЛИЗА
1.1.1 Роль микробиоты желудочно-кишечного тракта птицы
Целостность желудочно-кишечного тракта (ЖКТ) и микробного сообщества кишечника играют жизненно важную роль в усвоении пищи, развитии иммунитета и устойчивости к болезням. Изменения в микробном сообществе ЖКТ могут оказывать неблагоприятное воздействие на эффективность кормов, продуктивность и здоровье птиц (Jeurissen S. H., Lewis F., Van der Klis J. D., Mroz Z., Rebel J. M., Ter Huurne A. A., 2002; Kohl K. D., 2012). Понимание роли микробиоты ЖКТ цыплят и понимание современных методов, используемых
в исследованиях микробиома, имеет важное значение для улучшения микробиома ЖКТ птицы. Исторически сложилось так, что селективные культуральные методы использовались для идентификации и характеристики микробного разнообразия кишечника птиц. В последнее десятилетие использование секвенирования генов 16S рибосомной РНК (рРНК) значительно улучшило наше понимание состава и разнообразия микробиоты ЖКТ птиц, что привело к важным новым представлениям о биологической и экологической роли микробиоты ЖКТ. С помощью современных высокопроизводительных методов секвенирования можно быстро получить данные о полном видовом составе бактериального сообщества.
Желудочно-кишечные отделы птиц плотно заселены сложными микробными сообществами (бактериями, грибами, археями, простейшими и вирусами), в которых преобладают бактерии (Wei S., Morrison M., Yu Z., 2013). Взаимодействия между хозяином и бактериальным микробиомом куриного ЖКТ были широко изучены и рассмотрены многими исследовательскими группами (Danzeisen J. L., Kim H. B., Isaacson R. E., Tu Z. J., Johnson T. J., 2011; Hegde N. V., Kariyawasam S., DebRoy C., 2016; Kumar S. [et al.], 2018; Mancabelli L. [et al.], 2016; Pan D., Yu Z., 2014;) и в настоящее время считаются играющими важную роль в питании птиц, физиологии и развитии кишечника (Kau A. L., Ahern P. P., Griffin N. W., Goodman A. L., Gordon J. I., 2011).
Микробиота кишечника может образовывать защитный барьер, прикрепляясь к эпителиальным стенкам энтероцита, и тем самым снижая возможность колонизации патогенными бактериями (Yegani M., Korver D. R., 2008). Полезная микробиота продуцирует витамины (например, группы витамина К и B), короткоцепочечные жирные кислоты (уксусная кислота, масляная кислота
и пропионовая кислота), органические кислоты (например, молочная кислота)
и антимикробные соединения (например, бактериоцины), снижает уровень триглицеридов и индуцирует иммунные реакции, которые обеспечивают защиту животного (Jeurissen S. H., Lewis F., Van der Klis J. D., Mroz Z., Rebel J. M., Ter Huurne A. A., 2002; Yegani M., Korver D. R., 2008). С другой стороны, микробиом кишечника может также быть источником бактериальных патогенов, таких как Salmonella и Campylobacter, которые могут распространяться среди людей или выступать в качестве пула устойчивости к антибиотикам, т. е. представлять серьезную угрозу для здоровья населения (Zhou W., Wang Y., Lin J., 2012).
Нормальное микробное сообщество кишечника имеет и положительные,
и отрицательные стороны для хозяина (Dibner J. J., Richards J. D., 2005). Основные преимущества, которые обеспечивает комменсальная микробиота, – это конкурентное исключение патогенов или некоренных микробов (Dibner J. J., Richards J. D., 2005), иммунная стимуляция и программирование, а также вклад в питание хозяина. В более ранних работах было установлено, что условно выращенные животные гораздо менее восприимчивы к патогенам по сравнению
с животными, свободными от микробов (Koopman J. P. [et al.], 1984). Кроме того, комменсальная микробиота может стимулировать развитие иммунной системы, включая слой слизи, эпителиальный монослой, кишечные иммунные клетки (например, цитотоксические и хелперные Т-клетки, клетки, продуцирующие иммуноглобулин, и фагоцитарные клетки) и собственную пластинку – lamina propria (Dibner J. J., Richards J. D., 2005; Oakley B. B. [et al.], 2014; Shakouri M. D., Iji P. A., Mikkelsen L. L., Cowieson A. J., 2009). Эти ткани создают барьеры между хозяином и микробами и борются с нежелательными микроорганизмами кишечника. В дистальном отделе кишечника (т. е. в толстой кишке) микробиота также производит питательные вещества, такие как витамины, аминокислоты и короткоцепочечные жирные кислоты (SCFA) из непереваренного корма, которые в конечном итоге становятся доступными для хозяина (Dibner J. J., Richards J. D., 2005).
Также было установлено, что синтез SCFA снижает рН толстой кишки, что ингибирует превращение желчи во вторичные желчные продукты (Christl S. U., Bartram H. P., Paul A., Kelber E., Scheppach W., Kasper H., 1997). Кроме того, микробиота кишечника также способствует метаболизму азотистых соединений хозяина. Например, бактерии слепой кишки могут преобразовывать мочевую кислоту в аммиак, который впоследствии поглощается птицей и далее используется для производства аминокислот, таких как глутамин. Кроме того, часть азота из рациона включается в бактериальный клеточный белок, и поэтому сами бактерии могут быть источником белков/аминокислот.
С другой стороны, комменсальная микробиота также несет издержки для хозяина. В проксимальном отделе кишечника (желудке и тонком кишечнике) микробы конкурируют с хозяином за энергию и белок. Как в проксимальном, так и в дистальном отделе кишечника микробы продуцируют токсичные метаболиты (например, аминокислотные катаболиты) и катаболизируют желчные кислоты, которые могут подавлять рост и снижать усвояемость жира птицами. В присутствии микробиоты слизистый слой кишечника увеличивает секрецию муцина и скорость оборота эпителиальных клеток, тем самым сохраняя слизь желудочно-кишечного тракта и предотвращая проникновение микроорганизмов в эпителиальные клетки кишечника хозяина. Кишечная иммунная система также более развита и выделяет IgA, который специфически связывается с бактериальными эпитопами, помогает регулировать бактериальный состав в кишечнике (Suzuki K., Nakajima A., 2014). И хотя в целом эти процессы полезны, они увеличивают потребность в энергии и белке у хозяина и, следовательно, оказывают влияние на ростовые показатели птиц.
Дисбаланс микробиоты кишечника еще называют дисбактериозом. Он может быть определен как качественный и/или количественный дисбаланс нормальной микробиоты в тонком кишечнике, который может привести к последовательной реакции в ЖКТ, включая снижение барьерной функции кишечника (например, истончение кишечной стенки) и плохую усвояемость питательных веществ, и, следовательно, увеличение риска бактериальной транслокации
и воспалительных реакций (Yan F., Dibner J. J., Knight C. D., Vazquez-Anon M., 2017). Как неинфекционные, так и инфекционные стрессоры могут привести
к дисбактериозу. Неинфекционные факторы включают экологические стрессоры, пищевые дисбалансы, диетические изменения, микотоксины, ферментативную дисфункцию или генетику хозяина (Yan F., Dibner J. J., Knight C. D., Vazquez-Anon M., 2017). Инфекционные факторы включают вирусную или бактериальную инфекцию, кокцидиоз или токсические метаболиты, продуцируемые вредными микроорганизмами, такими как Clostridium perfringens.
Желудочно-кишечная микробиота может быть классифицирована как люминальная микробиота и слизистая (Jeurissen S. H., Lewis F., Van der Klis J. D., Mroz Z., Rebel J. M., Ter Huurne A. A., 2002). Состав люминальной микробиоты определяется наличием питательных веществ, наличием антимикробного вещества и скоростью прохождения корма. На состав микробиоты, прикрепленной
к слизистой оболочке, влияют несколько факторов хозяев, таких как экспрессия специфических участков адгезии на мембране энтероцитов, секреция секреторных иммуноглобулинов и скорость образования слизи. Люминальная микробиота и связанная со слизистой оболочкой микробиота, конечно, также влияют друг на друга (Jeurissen S. H., Lewis F., Van der Klis J. D., Mroz Z., Rebel J. M., Ter Huurne A. A., 2002), и поэтому важно признать, что диета может изменять как люминальную, так и присоединенную к слизистой оболочке микробиоту, чтобы влиять на здоровье кишечника. Насколько известно, до настоящего времени не было проведено ни одного исследования, в котором сравнивался бы таксономический состав или метаболические функции этих двух местообитаний микробов. Однако было бы интересно изучить и проанализировать различия между бактериальными сообществами слизистой оболочки на протяжении различных участков ЖКТ. Кроме того, изучение бактериального сообщества, связанного со слизистой оболочкой, важно для понимания реакции слизистой оболочки хозяина, поскольку любые изменения в иммунитете слизистой оболочки могут иметь серьезные последствия для здоровья птицы (Borda-Molina D., Vital M., Sommerfeld V., Rodehutscord M., Camarinha-Silva A., 2016).
1.1.2 Разнообразие микробиоты кишечника птицы
Желудочно-кишечный тракт птицы содержит разнообразное бактериальное сообщество, в котором каждая бактерия адаптирована к своей собственной экологической нише и синергически живет с другими бактериальными видами в том же сообществе. Показано, что состав и функции этих сообществ изменяются в зависимости от возраста птиц, расположения в желудочно-кишечном тракте и от компонентов питания (Kogut M. H., Oakley B. B., 2016).
Возраст птиц является одним из наиболее важных факторов, влияющих на бактериальный состав ЖКТ, плотность клеток и метаболическую функцию. Значительные изменения в таксономическом составе микробиоты кишечника были изучены с использованием методов DNA finger-printing (Lu J., Idris U., Harmon B., Hofacre C., Maurer J. J., Lee M. D., 2003) и высокопроизводительного секвенирования (Shaufi M. A., Sieo C. C., Chong C. W., Gan H. M., Ho Y. W., 2015), и хорошо изучены многими исследовательскими группами (Kers J. G., Velkers F. C., Fischer E. A. J., Hermes G. D. A., Stegeman J. A., Smidt H., 2018). Было продемонстрировано, что через один день после вылупления цыплята-бройлеры уже имеют микробное сообщество в их ЖКТ (Ballou A. L. [et al.], 2016; Kumar S. [et al.], 2018). Имеются также сукцессионные изменения в составе микробиома ЖКТ, обусловленные заменой и установлением более устойчивых бактериальных таксонов, по мере того как птица прогрессирует в возрасте (Crhanova M. [et al.], 2011; Lu J., Idris U., Harmon B., Hofacre C., Maurer J. J., Lee M. D., 2003). Лу с соавторами (Lu J., Idris U., Harmon B., Hofacre C., Maurer J. J., Lee M. D., 2003) обнаружили, что в ЖКТ цыплят в 3-дневном возрасте содержались L. delbrueckii, C. perfringens и Campylobacter coli, тогда как с 7 до 21-дневного возраста чаще встречались L. acidophilus, Enterococcus и Streptococcus. В 28-и и 49-дневном возрасте в желудочно-кишечном тракте содержится L. crispatus, но состав его значительно отличается от других возрастов (Lu J., Idris U., Harmon B., Hofacre C., Maurer J. J., Lee M. D., 2003). В другой работе сукцессионные изменения микробного сообщества кишечника, измеренные с помощью NGS, показали, что относительное обилие Clostridium было выше по мере старения птицы, тогда как лактобациллы были низкими на протяжении всего цикла роста. Такая вариабельность результатов может быть обусловлена типами проб (фекалии из слепой кишки) и/или традиционными микробиологическими и молекулярными методами, которые имеют ограниченный охват и точность по сравнению с высокопроизводительными платформами NGS, которые предлагают более высокий охват и глубину определения микробного сообщества. Технологии высокопроизводительного секвенирования, такие как целенаправленное ампликонное секвенирование и метагеномное секвенирование, стали более распространенными для анализа микробного состава и функций кишечника на протяжении всего жизненного цикла бройлеров, но все еще недостаточно знаний о морфологическом развитии хозяина и функциональных свойствах микробиома кишечника по мере старения птицы.
1.1.3 Строение желудочно-кишечного тракта птицы
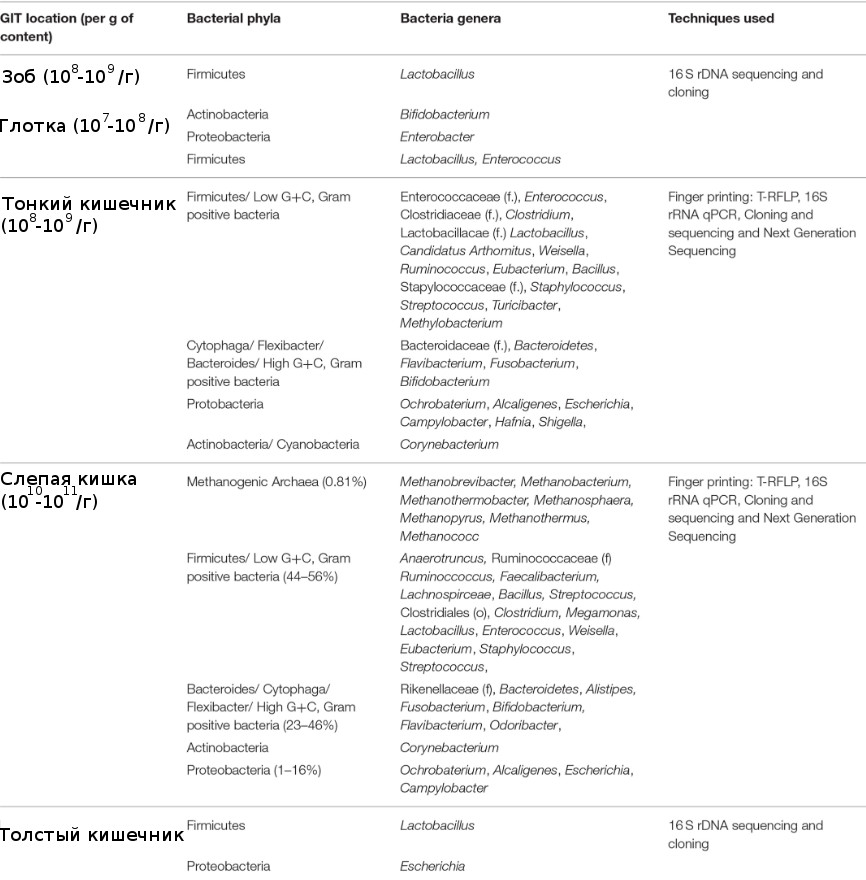
Желудочно-кишечный тракт цыпленка включает желудок, двенадцатиперстную кишку, тощую кишку, подвздошную кишку, толстую кишку и клоаку (Yeoman C. J., Chia N., Jeraldo P., Sipos M., Goldenfeld N. D., White B. A., 2012). Каждый участок желудочно-кишечного тракта имеет различные метаболические функции, которые формируют микробное сообщество (таблица 1), и поэтому, важно учитывать место отбора проб и дизайн исследования. В организме курицы, например, содержится от 108 до 109 КОЕ/г бактерий, в которых обычно преобладают лактобациллы (Gong J. [et al.], 2007).
В желудке концентрация бактерий низка, главным образом, из-за низкого pH, большинство бактерий в желудке – лактобациллы, энтерококки, лактоза-отрицательные энтеробактерии, и бактерии кишечной палочки (Gong J. [et al.], 2007). Среди сегментов тонкой кишки бактериальная плотность является самой низкой в двенадцатиперстной кишке из-за короткого времени прохождения
и разбавления секретируемой желчью (Waite D. W., Taylor M., 2015). Дуоденальное бактериальное сообщество в основном состоит из клостридий, стрептококков, энтеробактерий и лактобацилл (Waite D. W., Taylor M., 2015). Микробиота подвздошной кишки изучена больше всего среди сегментов тонкой кишки. Лу с соавторами (Lu J., Idris U., Harmon B., Hofacre C., Maurer J. J., Lee M. D., 2003) оценили бактериальный состав подвздошной кишки путем изучения последовательностей генов 16S рРНК и обнаружили больше всего содержание Lactobacillus (70 %), затем представители семейств Clostridiaceae (11 %), Streptococcus (6,5 %) и Enterococcus (6,5 %). По сравнению с подвздошной кишкой слепая кишка содержит более разнообразное, богатое и стабильное микробное сообщество, включая анаэробы (Videnska P., Sisak F., Havlickova H., Faldynova M., Rychlik I., 2013). Окли с соавторами (Oakley B. B. [et al.], 2014) задокументировали значительные изменения в микробных сообществах слепой кишки со дня вылупления до 6-недельного возраста у коммерческих бройлеров (Kogut M. H., Oakley B. B., 2016; Oakley B. B. [et al.], 2014), а также значительные различия в образцах слепой кишки и фекалий от одной особи (Kogut M. H., Oakley B. B., 2016). Как правило, богатство и разнообразие слепой кишки увеличивается в течение этих 6 нед, и таксономический состав сообщества быстро смещается от Proteobacteria, Bacteroides и Firmicutes, почти полностью Firmicutes к 3-недельному возрасту (Kogut M. H., Oakley B. B., 2016; Oakley B. B. [et al.], 2014). Однако Кумар с соавторами (Kumar S. [et al.], 2018) обнаружили, что фирмикуты были наиболее распространенным типом как в толстой кишке, так и в подвздошной кишке во всех возрастах (день 0 – день 42). Различия в бактериальном составе можно объяснить различиями в протоколе экстракции нуклеиновых кислот, праймерах, подходе к секвенированию, факторах окружающей среды, диетическом лечении/составе, породе и географических условиях. В дополнение к типам образцов, адекватный размер выборки также необходим для правильного дизайна исследования. Более высокие индивидуальные различия в типах образцов (образцы культур) приводят к более высокому размеру выборки по сравнению с образцами слепой кишки, чтобы найти потенциальные различия (Lagkouvardos I., Overmann J., Clavel T., 2017).
Таблица 1 – Распределение наиболее распространенных бактериальных
таксонов в ЖКТ птиц независимо от возраста, рациона питания
(Chicken Gut Microbiota: Importance and Detection Technology, 2018)
Также известно, что кормовые компоненты и добавки, подходы к обработке кормов оказывают влияние на микробное сообщество кишечника Кнареборг с соавторами (Knarreborg A., Simon M. A., Engberg R. M., Jensen B. B., Tannock G. W., 2002) заявляют, что измельченный корм снижает количество Enterococcus spp.и кишечной палочки, но увеличивает содержание Lactobacillus spp. и C. perfringens в подвздошной кишке бройлеров по сравнению с гранулированным кормом (Knarreborg A., Simon M. A., Engberg R. M., Jensen B. B., Tannock G. W., 2002). Кумар с соавторами (Kumar S. [et al.], 2018) сообщили
о низкой численности Firmicutes и высокой численности Bacteroidetes с 0-го по 42-й день, когда птицы перешли от стартовой диеты к финишной, и утверждали, что представители типа Bacteroidetes жизненно важны для ферментации крахмала до простых сахаров. Кроме того, кормовые добавки, такие как сбраживаемые сахара (пребиотики), также могут оказывать влияние на состав
и разнообразие микробиоты кишечника цыпленка.
1.1.4 Исследование микробиоты из кишечника сельскохозяйственной птицы с помощью молекулярных подходов
Классические методы, основанные на изучении бактериальных культур, исторически широко использовались для изучения микробиоты кишечника. Тем не менее, эти методы очень избирательны к культивируемым бактериям
в определенных условиях. Большинство бактерий являются некультультивируемыми. Более 10 лет назад было замечено, что из 52 известных типов микроорганизмов только половина имела хотя бы одного культивируемого представителя, и это подтверждает описание «некультивируемого большинства» (Rappe M. S., Giovannoni S. J., 2003). Следовательно, представленность (количество видов) и разнообразие (количество видов, взвешенных по их относительной численности) кишечных бактерий были недооценены, и наши знания о кишечной микробиоте остаются неполными (Gong J. [et al.], 2002).
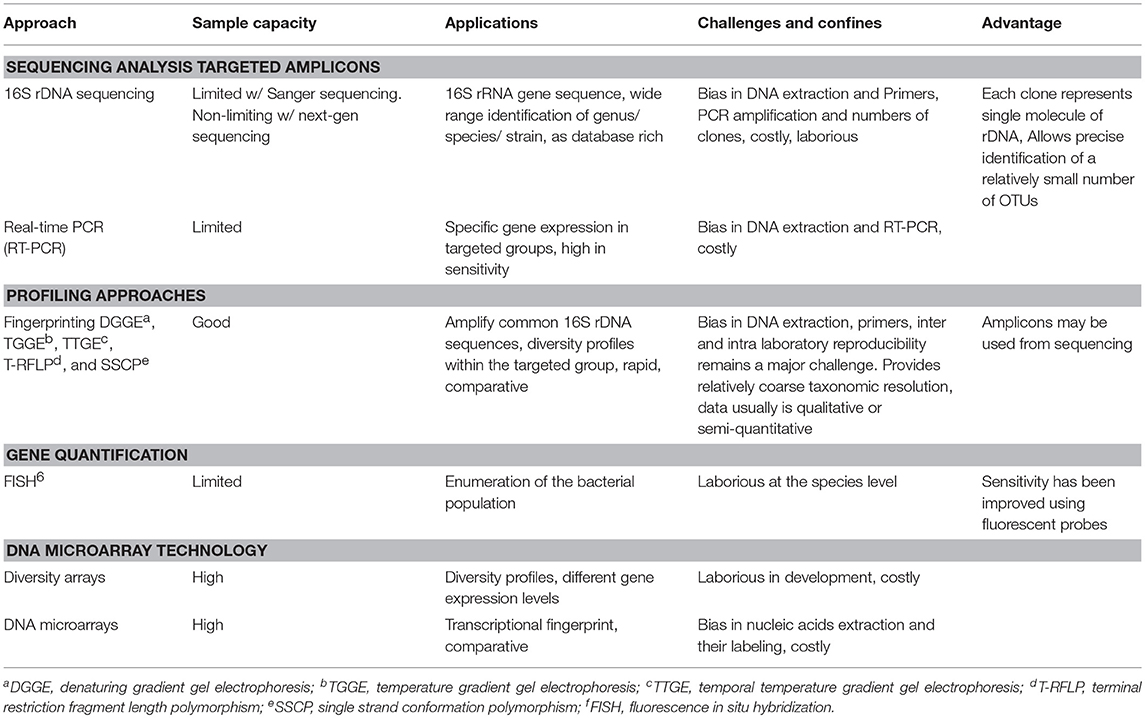
Развитие молекулярной биотехнологии предложило новые инструменты для изучения состава, разнообразия, прогнозируемой функции и взаимодействия кишечной микробиоты в различных отделах желудочно-кишечного тракта. В настоящее время доступны различные молекулярные методы, каждый из которых имеет свои сильные и слабые стороны. В таблице 2 представлены состав образца, применение и ограничения некоторых из применяемых наиболее распространенных молекулярных методов, которые можно использовать для изучения экологических микробных микроорганизмов микробиома сельскохозяйственной птицы. Высокопроизводительное секвенирование ампликонов гена 16S рРНК быстро стало наиболее применяемым методом. Хотя этот метод широко использовался в других областях исследований, в 2013 г. был опубликован первый отчет, в котором было использовано высокопроизводительное секвенирование генов 16S рРНК для изучения популяции микробных сообществ и их взаимодействий в курином ЖКТ (Diaz-Sanchez S., Hanning I., Pendleton S., D’souza D., 2013).
Таблица 2 – Молекулярные подходы на основе 16S рРНК для изучения
микробиома ЖКТ курицы. Цитировано по Chicken Gut Microbiota: Importance and Detection Technology, 2018
Молекула 16S рРНК представляет собой небольшую субъединицу рибосомы, которая содержит последовательности, которые высоко консервативны для всех бактерий. Для амплификации этих генов метагеномную ДНК выделяют из образцов фекалий или содержимого ЖКТ, и амплифицируют методом полимеразной цепной реакции (ПЦР) с помощью праймеров на консервативные участки гена 16S рРНК (Apajalahti J., Kettunen A., Graham H., 2004). Секвенирование этих амплифицированных продуктов (ампликонов) позволяет различить бактерии, как правило, на уровне рода или вида (Flint H. J. [et al.], 2006), а относительная численность каждой последовательности отражает относительную численность этой бактерии в исходном образце. Таким образом, секвенирование генов 16S рРНК обеспечивает реальный состав бактериального сообщества
с помощью определения типов бактерий, присутствующих в образце, и их относительной численности. Из-за высокой представленности и разнообразия кишечных бактериальных сообществ только в последние несколько лет технология секвенирования ДНК достигла такого уровня, что теперь можно полностью определить количественный состав метагеномных сообществ. Начиная
с 2008 г., технический прогресс в секвенировании позволил анализировать на несколько порядков больше последовательностей, чем это было возможно ранее (Hamady M., Walker J. J., Harris J. K., Gold N. J., Knight R., 2008). Благодаря этим глубоким методологическим достижениям и огромным новым базам данных, теперь можно легко и точно провести анализ микробиома ЖКТ птицы, чтобы определить, например, как микробиом реагирует на различные кормовые добавки, условия содержания или болезненные состояния (рисунок 1).
Высокопроизводительное секвенирование, или секвенирование следующего поколения (NGS), является мощным инструментом для исследования биологической и экологической роли кишечной микробиоты (Diaz-Sanchez S., Hanning I., Pendleton S., D’souza D., 2013). NGS стал удобным, быстрым, точным и недорогим методом геномных исследований (Park S. H., Hanning I., Perrota A., Bench B. J., Alm E., Ricke S. C., 2013). Современные платформы NGS обеспечивают высокую пропускную способность, быстрое время исследования и невысокие затраты. Среди этих платформ приборы Illumina HiSeq и MiSeq являются двумя наиболее часто используемыми системами в недавних исследованиях
в области микробиома и метагеномики в ЖКТ. Несмотря на многие преимущества, эти платформы имеют ограничения, например, краткую сборку прочтения и высокую стоимость (Kumar S., Pitta D. W., 2015). Платформы секвенирования третьего поколения, такие как single molecule real-time (SMRT) и секвенирование нанопор, требуют меньше времени для подготовки ДНК (без ПЦР) и являются экономически эффективными (Kumar S., Pitta D. W., 2015). Поскольку эти платформы продолжают развиваться, их применение, несомненно, приведет
к новому пониманию микробиома желудочно-кишечного тракта птицы.
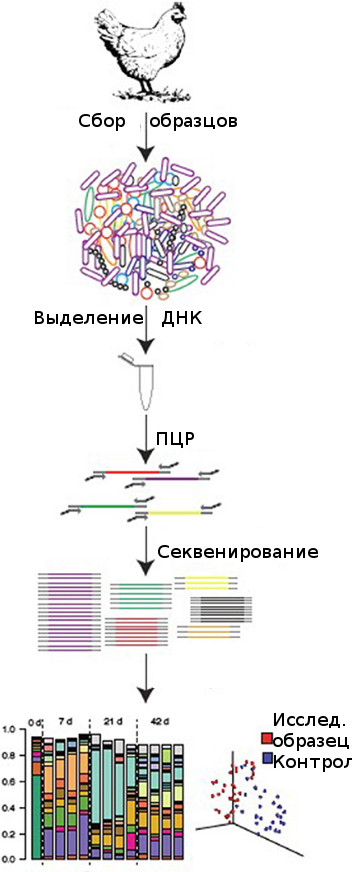 |
Рисунок 1 – Стандартная процедура исследования микробиома птицы |
Для биоинформационного анализа данных последовательностей, полученных в результате секвенирования, требуются платформы с открытым исходным кодом, такие как QIIME или Mothur, которые используют общедоступные базы данных GreenGenes, проект базы данных Ribosomal и SILVA (Caporaso J. G.
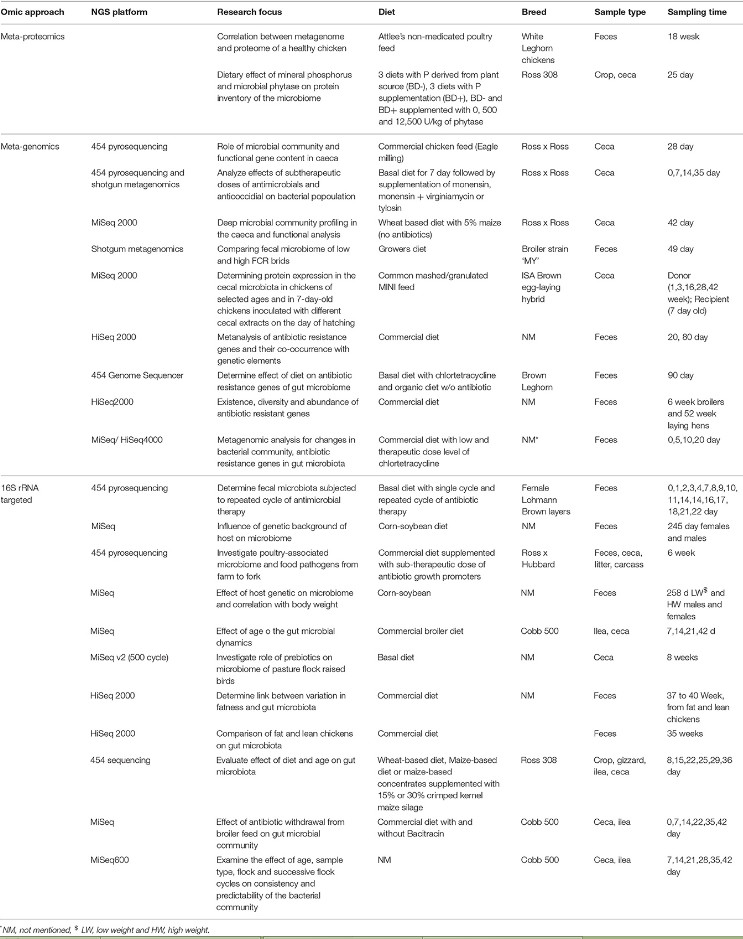
[et al.], 2010) для выполнения таксономического ранжирования. Предсказание метаболических функций на основе таксономических идентификаторов последовательностей генов 16S рРНК может быть дополнительно получено с использованием таких алгоритмов, как PICRUSt и Tax4Fun (Abhauer K. P., Wemheuer B., Daniel R., Meinicke P., 2015). Для анализа функций генов или отдельных геномов могут использоваться метагеномный или метатранскриптомный подходы (в которых гены или транскрипты, соответственно, секвенируются напрямую без ПЦР) для предоставления информации о разнообразии сообществ, структуре и метаболических функциях или экспрессии генов. Биоинформационный анализ таких наборов данных является более сложным, чем анализ амплификации 16S, и обычно содержит сборщик последовательностей, такой как Velvet (CLC workbench, Newbler version 3.0, Biospace) или MG-RAST. Бактериальные таксоны и функциональные группы могут быть определены на основе Basic Local Alignment Инструмент поиска (BLAST), а функции генов могут быть проанализированы с использованием либо Киотской энциклопедии генов и геномов (KEGG), либо кластера ортологичных генов (COG). Например, для изучения микробиома ЖКТ птиц с целью выяснения функций слепой кишки и реакции кишечника на заражение патогенами, а именно изучение корреляции между представленностью микробиологического сообщества и параметрами производительности, выявление генов устойчивости к антибиотикам и др., проводили метагеномные исследования (Borda-Molina D., Seifert J., Camarinha-Silva A., 2018). Некоторые из исследований, основанных на NGS, в которых изучаются состав и функции микробного сообщества кишечника цыпленка в отношении изменения рациона/лечения антибиотиками, представлены в таблице 3. Однако для изучения микробиома кишечника и получения сопоставимых результатов необходим стандартный протокол. В настоящее время большинство метагеномных подходов к изучению куриного ЖКТ до сих пор недоступны для большинства исследователей или ветеринаров.
Таблица 3 – Применение разных омических подходов для изучения
микробного сообщества и функций кишечника птиц
Чтобы обойти некоторые ограничения анализа методом NGS, для определения метаболических и функциональных свойств микробиома недавно были также использованы протеомные подходы (Tilocca B., Witzig M., Rodehutscord M., Seifert J., 2016). Транскриптомика измеряет транскрипцию генов in situ, обеспечивая точное отражение физиологических функций (Kumar S., Pitta D. W., 2015). Поскольку в современных базах данных количество опубликованных бактериальных штаммов ЖКТ птицы ограничено, увеличение количества бактериальных культур и надлежащая каталогизация их биохимических и генетических свойств облегчит протеомику и другие «омические» подходы.
1.1.5 Пребиотики и пробиотики в рационе сельскохозяйственной птицы
Использование пребиотиков в качестве пищевых модуляторов оказывает положительное влияние на некоторые бактериальные таксоны в толстой кишке (Gaggia F., Mattarelli P., Biavati B., 2010). Например, фруктоолигосахариды (FOS) и галактоолигосахариды (GOS) увеличивают популяцию Bifidobacterium и Lactobacillus (Jung S. J., Houde R., Baurhoo B., Zhao X., Lee B. H., 2008; Xu Z. R., Hu C. H., Xia M. S., Zhan X. A., Wang M. Q., 2003). Исследования in vitro показали, что фекальные взвеси, которые инкубировали с олигофруктозой и инулином, демонстрировали увеличение популяций бифидобактерий в толстой кишке человека, тогда как потенциальные патогены, такие как Escherichia coli
и Clostridium spp. поддерживались на более низких уровнях. Большинство штаммов бифидобактерий (например, B. fiagilk, B. thetaiotaomicron, B. vulgatus, B. dktasonk и B. ovatus), за исключением B. bifidum, могут использовать FOS
в качестве стимулятора роста и ферментации. Эти бактерии выделяют фермент бета-фруктозидазу, которая может легко перерабатывать и ферментировать FOS. Однако микроорганизмы, такие как E.coli и C. perfringens, не способны использовать FOS в качестве источника углевода. Было показано, что у крыс, которых кормили FOS, временно увеличивалось содержание бактерий, продуцирующих молочную кислоту, и долгосрочно повышался уровень масляной кислоты в слепой кишке. Включение в рацион бройлеров FOS уменьшало популяции C. perfringens и E. coli и увеличивало разнообразие Lactobacillus
в ЖКТ бройлеров (Kim H. J., Eom S. J., Park S. J., Cha C. J., Kim G. B., 2011). Паттерсон с соавторами (Patterson J. A., Orban J. I., Sutton A. L., Richards G. N., 1997) оценивали влияние термических кетозных олигосахаридов на микробные популяции слепой кишки цыплят бройлеров. Результаты показали, что концентрации бифидобактерий и лактобацилл слепой кишки были увеличены в 24 раза и в 7 раз, соответственно, при добавлении в рацион кетоз по сравнению с контрольной группой.
Другой тип пребиотиков – манноолигосахариды (MOS) – имеет различные механизмы действия (Patterson J. A., Orban J. I., Sutton A. L., Richards G. N., 1997). Манноолигосахариды выполняют несколько функций:
1) связываются с потенциальными патогенными грамотрицательными бактериями (например, E. coli и Salmonella), которые обладают фимбриями 1-го типа (чувствительными к маннозе лектином), для предотвращения прикрепления патогенов к стенке кишечника;
2) проявляют иммуномодулирующий эффект, основанный на особенностях антигенности компонентов маннана и глюкана;
3) модулируют морфологию кишечника;
4) усиливают экспрессию муцина и снижают скорость энтероцитов (Yang Y., Iji P. A., Choct M., 2009).
Воздействие пребиотиков на нижний отдел желудочно-кишечного тракта включает:
1) подачу и ферментацию пищи для микробиоты слепой и толстой кишок;
2) производство конечных продуктов ферментации (например, SCFA);
3) стимуляцию сахаролитической ферментации;
4) подкисление содержимого толстой кишки;
5) гиперплазию эпителия слепой и толстой кишок;
6) стимуляцию секреции гормональных пептидов толстой кишки (Gaggia F., Mattarelli P., Biavati B., 2010).
Состав кишечной микробиоты помимо возраста, места ЖКТ и пребиотиков во многом зависит от породы и пола птицы (Kers J. G., Velkers F. C., Fischer E. A. J., Hermes G. D. A., Stegeman J. A., Smidt H., 2018). Кроме того, было показано, что факторы окружающей среды (уровень биологической безопасности, жилье, мусор, доступ к корму и климат) также существенно влияют на бактериальный состав кишечника. Таким образом, интерпретация данных и результаты исследований во многом зависят от дизайна исследования.
Наряду с пребиотиками, пробиотики являются инструментом, используемым для уменьшения применения противомикробных препаратов в сельском хозяйстве (Borda-Molina D., Seifert J., Camarinha-Silva A., 2018). Пробиотики являются жизнеспособными бактериями, которые обеспечивают пользу при приеме внутрь, в том числе усиление функций кишечного барьера хозяина против потенциальных патогенов, поддержание гомеостаза в ЖКТ (Bajagai Y. S., Klieve A. V., Dart P. J., Bryden W. L., 2016). Пробиотики могут принести пользу хозяину напрямую без изменения микробиоты в целом (Bajagai Y. S., Klieve A. V., Dart P. J., Bryden W. L., 2016). У самцов бройлеров добавление в рацион Bacillus subtilis CGMCC 1.1086 привело к увеличению прироста веса и улучшению коэффициента конверсии корма (FCR) (Li Y., Xu Q., Huang Z., Lv L., Liu X., Yin C., Yan H., Yuan J., 2016). Другое исследование включало кормление пробиотиком Lactobacillus planatarum, в результате чего повышался иммунитет, наблюдали увеличение секреции сывороточного IgG и IgA (Gao P., Ma C., Sun Z., Wang L., Huang S., Su X., Xu J., Zhang H., 2017). В качестве пробиотиков у домашней птицы обычно применяются штаммы Lactobacillus и Enterococcus spp., оба обнаружены в естественной микробиоте из ЖКТ птицы в большом количестве (Markowiak P., Slizewska K., 2018; Pourabedin M., Guan L., Zhao X., 2015). Добавление Lactobacillus spp. в рацион связывают с увеличением массы тела, увеличением количества бокаловидных клеток и снижением колонизации кишечной палочки в пищеварительном тракте (Ashraf S., Zaneb H., Yousaf M. S., Ijaz A., Sohail M. U., Muti S., Usman M. M., Ijaz S., Rehman H., 2013). Добавка в пищу Enterococcus spp. повышает коэффициент конверсии корма (FCR) и положительно влияет на рост бройлеров (Samli H. E., Senkoylu N., Koc F., Kanter M., Agma A., 2007). Stanley с соавторами продемонстрировали, что представители семейств Lachnospiraceae, Ruminococcaceae и Erysipelotrichaceae являются потенциальными пробиотиками, их добавление в рацион птицы значительно коррелировало с хорошими показателями коэффициент конверсии корма (Stanley D., Hughes R. J., Geier M. S., Moore R. J., 2016). В дугой работе Torok с соавторами показали ассоциацию применения Lactobacillus spp. с плохой продуктивностью животных (Torok V. A., Hughes R. J., Mikkelsen L. L., Perez-Maldonado R., Balding K., MacAlpine R., Percy N. J., Ophel-Keller K., 2011). Кстати, оба исследования назвали Clostridium lactatifermentans потенциальным пробиотиком для выращивания сельскохозяйственных птиц.
В большинстве случаев добавление в рацион только пребиотиков или пробиотиков не всегда приводит к желаемому эффекту. Даже после прохождения полного курса нормализации баланса микроорганизмов в ЖКТ, уже через две недели–месяц пробиотические микроорганизмы погибают, и уровень микроорганизмов становится прежним. В связи с этим образовалась стратегия создания нового средства коррекции микрофлоры желудочно-кишечного тракта с эффектом синергизма в результате комбинированного действия про- и пребиотиков (Alloui M. N., Szczurek W., Swiatkiewicz S., 2013).
Синбиотики – средства нового поколения для нормализации микрофлоры кишечника, которые включают пробиотики и пребиотики. Первые представляют собой бактерии, которые положительно влияют на здоровье человека. Они усиливают барьерные функции слизистой оболочки кишечника, восстанавливают ее целостность, улучшают работу иммунной системы, метаболизм, продуцируют витамины группы В. Вторые, в свою очередь, – не перевариваемые углеводы, стимулирующие рост микроорганизмов-пробиотиков. Только за последнее время проведено огромное количество исследований, показывающих положительное влияние синбиотических добавок на рост, вес, снижение значения FCR и увеличение сохранности птицы, но ни один из показателей не является двукратным повышением эффективности применения синбиотиков по сравнению с использованием только про- или пребиотических добавок (Awad W. A., Ghareeb K., Abdel-Raheem S., Bohm J., 2009; Mohnl M., Acosta Aragon Y., Acosta Ojeda A., Rodriguez Sanchez B., Pasteiner S., 2007; Mookiah S., Sieo C. C., Ramasamy K., Abdullah N., Ho Y. W., 2014). Определено влияние синбиотических композиций на слизистую кишечника (сохранение рисунка ворсинок, глубины криптов) (Sohail M. U., Hume M. E., Byrd J. A., Nisbet D. J., Ijaz A., Shabbir M. Z., Rehman H., 2012). Принимая во внимание теоретически возможные синергические эффекты альтернативных добавок этого типа, потенциал использования синбиотиков в сельском хозяйстве никто не отвергает, но дальнейшее развитие этого подхода в птицеводстве пока не получает нужного внимания (Sohail M. U., Hume M. E., Byrd J. A., Nisbet D. J., Ijaz A., Shabbir M. Z., Rehman H., 2012).
1.1.6 Компоненты ветеренарных препаратов и кормовых добавок (органические кислоты, ферменты, антимикробные пептиды)
Органические кислоты
В качестве пищевых добавок в птицеводстве применяются карбоновые (муравьиная, уксусная, пропионовая, масляная) и оксикарбоновые (молочная, яблочная, винная, лимонная) кислоты (Dibner J. J., Buttin P., 2002). Эти вещества широко распространены в природе и являются метаболитами процессов усвоения пищи (биологического окисления жиров, сахаров, белков) у живых организмов.
Препараты на основе органических кислот и/или их солей применяются
в виде отдельных водных растворов или коктейлей (при выпаивании) или в виде сухих кормовых добавок в различных рационах (Huyghebaert G., Ducatelle R., Van Immerseel F., 2011).
Выгода от использования органических кислот как альтернативных добавок известна уже давно. Еще в 1988 г. было показано, что добавление фумаровой кислоты в рацион цыплят увеличивало такие производственные показатели как привес и усвояемость корма (Patten J. D., Waldroup P. W., 1988). Подобное влияние на производственные показатели роста продукции птицеводства были продемонстрированы для масляной, молочной и других кислот (Adil S., Banday T., Bhat G. A., Salahuddin M., Raquib M., Shanaz S., 2011; Hernandez F., Madrid J., Garcia V., Orendo J., Megias M. D., 2006; Salgado-Transito L., Rio-Garcia J. C., Moreno-Martinez J. L., Mendez-Albores A., 2011). Подробно исследовано влияние смесей органических кислот в составе коктейлей (Samanta S., Haldar S., Ghost T. K., 2010).
Механизм влияния добавок органических кислот еще не изучен. Но повышение эффективности потребления пищи у птиц в случае использования органических кислот связывается, в первую очередь, с антимикробными свойствами этих соединений. Механизмы влияния могут быть взаимосвязаны: 1) с понижением значений рН в верхних отделах ЖКТ птиц (зоб, железистый желудок, мускульный желудок) и изменениями физиологических свойств мукозального слоя (Panda A. K., Rama S. V., Raju M. N., Shyam Sunder G., 2009); 2) изменениями состава микрофлоры за счет уменьшения количества кислотолабильных патогенных бактерий и повышения числа автохтонной кислотоустойчивой молочно-кислой микрофлоры, обладающей пробиотическими свойствами (Boroоjeni F., Vahjen W., Mader A., Knorr F., Ruhnke I., Rohe I., Hafees A., Villodre C., Manner K., Zentek J., 2014); 3) повышением усвояемости пищи, за счет повышения активности метаболизирующих энзимов и перехода витаминов и минеральных веществ из малодоступных или труднодоступных для организма соединений в более доступные формы (Nezhad Y. E., Gale-Kandi J. G., Farahvash T., Yeganeh A. R., 2011); 4) регенерационным потенциалом эпителиальных клеток, связанным
с секретами и/или продуктами метаболизма симбионтной микрофлоры. Влияние препаратов органических кислот на повышение рентабельности птицеводства при их вводе в рационы бройлеров очевидно, но из-за многообразия видов, форм и методов использования научные результаты исследования механизмов влияния этих соединений пока не показаны (Kim J. W., Kim J. H., Kil D. Y., 2015).
Ферменты
Общепринятые способы производства комбикормов с использованием
в рационах исключительно животного белка в настоящее время не могут удовлетворить растущие потребности промышленного птицеводства. Методом использования растительного сырья и применения биотехнологических приемов его переработки в настоящее время создаются новые функциональные продукты для кормления птицы.
Основными источниками углеводов для птицы остаются зерновые корма. Они неоднородны по своему составу. Зерновые корма (кукуруза, пшеница, ячмень, овес) имеют в своем составе практически одинаковое содержание углеводов, но качественные и количественные различия содержания олигосахаридов, декстринов, крахмала, целлюлозы, лигнина влияют на перевариваемость корма и, как следствие, его использования в организме.
Организм птицы способен вырабатывать ферменты, гидролизующие почти все крупные молекулы корма на мономеры для последующего усвоения в организме, но в силу физиологических причин качество работы собственной ферментной системы может быть недостаточным из-за избыточного присутствия
в современных рационах трудногидролизуемых компонентов, ингибиторов протеолитических ферментов и ряда других, менее значимых для питания компонентов. Поэтому применение ферментных препаратов в кормопроизводстве продиктовано необходимостью уменьшения или полным исключением влияния содержащихся в сырье антипитательных веществ на биологическую ценность рациона.
В качестве кормовых добавок традиционно в птицеводстве используются три группы ферментов: фитазы, протеазы, карбогидразы (ксиланазы, целлюлазы, галактозидазы, манназы, амилазы и пектиназы).
Ферменты, используемые в промышленных масштабах, обычно имеют микробное или грибковое происхождение. С момента первых публикаций
Ф. Кликнера и Е. Фолуэлла (1926 г., цитировано по Швыдков А. Н., Мартышенко А. Е., Ланцева Н. Н.,. Чебаков В. П, Кобцева Л. А., 2014) об улучшении роста цыплят (и повышения яйценоскости кур за счет применения в комбикормах протезима) эффективное влияние ферментных добавок на такие производственные показатели как повышение продуктивности птицы и усвояемость корма доказана многочисленными обзорами, например (Adeola O., Cowieson A. J., 2011; Bedford M. R., Schulze H., 1998; Woyengo T. A., Nyachoti C. M., 2011). Поэтому для улучшения перевариваемости и питательности кормов
в настоящее время ферментные добавки кормов в рационах животных используются повсеместно.
Помимо прямого улучшения гидролизуемости растительных добавок,
и, как следствие, усвояемости корма, введение карбогидраз в промышленные рационы опосредованно может влиять и на состав микрофлоры ЖКТ птицы. Как было уже сказано выше, растительные полисахариды обладают пребиотическими эффектами, поскольку после ферментации они могут являться питательными веществами полезной микрофлоры. Так ферменты могут индуцировать заселение ЖКТ конкурентоспособными штаммами-пробионтами и осуществлять контроль над численностью условно-патогенной микрофлоры (Cheng G., Hao H., Xie S., Wang X., Dai M., Huang L., Yuan Z., 2014).
Дороговизна в обеспечении бройлеров кормами животного происхождения вызывает постоянную необходимость разработки кормов растительного типа
и применения биотехнологических приемов его ресурсосберегающей переработки. Из-за этого в условиях промышленного птицеводства обогащение кормов ферментными препаратами на сегодня является обязательным условием.
И роль ферментных (полиферментных) добавок для повышения перевариваемости и питательности кормов огромна. Например, введение в рецептуру рационов ферментных добавок препаратов фитазы в сочетании с β-манназой может приводить к увеличению веса бройлеров на 4,2 %, и к уменьшению такого показателя как FCR вплоть до 4,8 %.
Однако постоянно следует иметь ввиду множественные расхождения
в количественных показателях эффективности использования комплексных ферментных добавок, зависящих от вида и источника белка, соотношения входящих в рацион бройлера компонентов, побочных реакций, генетических особенностей кросса и многих других (Cheng G., Hao H., Xie S., Wang X., Dai M., Huang L., Yuan Z., 2014).
Антимикробные пептиды
К числу антимикробных препаратов относятся и эндогенные антимикробные пептиды (АМП). Они представляют собой рибосомально синтезируемые молекулы и включают в себя 10–100 аминокислотных остатков. АМП различаются большим содержанием гидрофобных основных остатков лизина и аргинина, обладают амфифильными свойствами и проявляют активность против широко круга патогенов. Механизм прямого антимикробного действия, как правило, связан с нарушением целостности мембраны мишени, однако также и опосредован специфическим взаимодействием с поверхностными или внутриклеточными мишенями.
На сегодняшний день известно уже более 2600 представителей семейства АМП и их рекомбинантных аналогов (http://aps.unmc.edu/AP/main.php; (Fosgerau K., Hoffmann T., 2015)). Их условно можно разделить на три группы: а) цистеин-содержащие пептиды, стабилизированные внутримолекулярными дисульфидными связями; б) линейные пептиды, содержащие в своем составе
α-спиральные участки и в)линейные пептиды, обогащенные остатками определенных аминокислот.
Поскольку выработка популяционной устойчивости у бактерий к АМП занимает длительное время, с биологической точки зрения АМП являются эффективной альтернативой антибиотикам микробного происхождения. Поэтому большинство исследований связано с изучением действия АМП против патогенов, вызывающих инфекционные заболевания.
Однако в некоторых исследованиях были продемонстрированы промоторные ростовые свойства АМП и показано, что добавление АМП в корм продуктивных животных улучшает их сохранность, нормализует микрофлору
и морфологию ЖКТ. Одним из примеров такого действия, т. е. использования АМП как альтернативной добавки, было применение в рационах промышленной птицы генноинженерного аналога цекропина А (1-11)-D-(12-37)-Asn-(CADN). Подобные результаты были также получены с еще одним химерным вариантом этого же класса пептидов – магаинином (Choi S. C., Ingale S. L., Kim J. S., Park Y. K., Kwon I. K., Chae B. J., 2013).
Иммуностимулирующая активность АМП была доказана группой ученых, которые на протяжении нескольких лет оценивали изменение роста промышленной птицы при ее выпаивании экстрактами лимфоидного органа желудков (Sacculus rotundus) кроликов и свиней (Wang D., Ma W., She R., Sun Q., Liu Y., Hu Y., Liu L., Yang Y., Peng K., 2009). При этом было показано, что использование природных пептидов в качестве кормовой добавки улучшает рост птиц
и сопровождается увеличением количества интраэпителиальных Т-лимфоцитов (популяция, попадающая из периферической крови в прослойку между базолатеральными мембранами эпителиальных клеток, играет важную роль
в иммунной защите, осуществляемой слизистой тонкой кишки), тучных клеток и уровня IgA.
АМП, продуцируемые бактериями и угнетающие развитие родственных микроорганизмов получили название «бактериоцины».
1.2 МЕТОДЫ ПОЛНОГЕНОМНОГО СЕКВЕНИРОВАНИЯ БАКТЕРИЙ
Секвенирование целого генома является важным инструментом для картирования геномов новых организмов, разделения геномов известных организмов или сравнения геномов по нескольким образцам. Секвенирование целых бактериальных, вирусных и других микробных геномов важно для создания точных эталонных геномов, для идентификации и для других сравнительных геномных исследований.
В отличие от капиллярного секвенирования или подходов, основанных на ПЦР, секвенирование следующего поколения (NGS) позволяет исследователям секвенировать сотни организмов. В отличие от традиционных методов секвенирование микробного генома на основе NGS не зависит от трудоемких этапов клонирования, экономя время и упрощая рабочий процесс. NGS может идентифицировать низкочастотные варианты и перестройки генома, которые могут быть пропущены или являются слишком трудоемкими для идентификации
с использованием других методов.
Полногеномное секвенирование бактерий de novo включает сборку генома без использования геномного референса, и часто используется для секвенирования новых микробных геномов. Современные секвенаторы обеспечивают точность считывания, длину и глубину считывания для получения высококачественных драфтов и полных сборок микробного генома.
Полногеномное ресеквенирование включает секвенирование всего генома бактерии, вируса или другого микроба и сравнение последовательности с известным драфтом. Генерирование быстрой и точной информации о последовательности микробного генома имеет решающее значение для выявления низкочастотных мутаций, поиска ключевых делеций и вставок, а также для обнаружения других генетических изменений среди штаммов микроорганизмов.
Система MiSeq (Illumina) наиболее удобная система для проведения целенаправленного ресеквенирования и секвенирования генома малого размера (www.illumina.com/miseq).
Основные особенности системы MiSeq:
• Исключительное качество данных, продемонстрированное по результатам рецензируемого научного сравнения.
• Прибор с простым и интуитивно-понятным рабочим процессом.
• Наименьшее время выполнения цикла. Самое быстрое секвенирование
и обнаружение вариантов позволяет проводить срочные исследования.
• Обширный набор областей применения. Настраиваемая длина считывания и различные типы проточных кювет обеспечивают максимальную адаптируемость к широкому диапазону областей применения.
Система MiSeq предлагает первое комплексное решение для секвенирования, объединяя в одном приборе генерацию кластеров, амплификацию, секвенирование и анализ данных. Благодаря небольшой площади занимаемой поверхности – приблизительно 0,2 м2 – система с легкостью вписывается буквально в любую лабораторную среду. В системе MiSeq используется секвенирование посредством технологии синтеза (SBS) от компании Illumina, наиболее широко используемой химической реакции секвенирования нового поколения.
На сегодняшний день существует более 750 публикаций о системе MiSeq, которая является идеальной платформой для проведения быстрого и экономичного генетического анализа. Исключительное качество данных. Химическая реакция на основе SBS от компании Illumina – это технология секвенирования нового поколения, которая получила наиболее широкое распространение. Исключительное качество данных достигается благодаря химической реакции на основе технологии SBS – запатентованного обратимого метода на основе нуклеотидов, прерывающих цепь, который выявляет одиночные основания по мере их встраивания в массивно параллельные цепи ДНК. Флуоресцентные красители нуклеотида, прерывающего цепь, визуализируются по мере добавления дНТФ с последующим расщеплением, что обеспечивает встраивание следующего основания. Благодаря наличию в каждом цикле всех четырех дНТФ, обратимо связанных с нуклеотидом, прерывающим цепь, естественная конкуренция сводит к минимуму систематическую ошибку встраивания. Распознавания оснований производятся непосредственно по измерениям интенсивности сигнала во время каждого цикла, что значительно снижает частоту возникновения серьезных ошибок по сравнению с другими технологиями (Junemann S. [et al.], 2013; Liu L. [et al.], 2012; Loman N. J. [et al.], 2012; Quail M. A. [et al.], 2012; Ravi R. K., Walton K., Khosroheidari M., 2018; Ross M. G. [et al.], 2013). Результатом является последовательное секвенирование оснований с высокой точностью, что фактически устраняет ошибки последовательности, зависящие от контекста, даже в пределах участков с повторяющейся последовательностью. Высочайшая производительность секвенирования Illumina обеспечивает получение данных без ошибок при проведении анализа наиболее чувствительных или сложных образцов.
Прибор с простым и интуитивнопонятным рабочим процессом. Управляющее программное обеспечение прибора системы MiSeq является простым
и понятным. Интуитивно-понятный интерфейс на основе сенсорного экрана, автоматически настраиваемые картриджи с реактивами с функцией отслеживания RFID, возможность просматривать обучающие видео и следовать пошаговым инструкциям в каждом рабочем процессе секвенирования. Все системы MiSeq имеют возможность автономного проведения анализа данных и доступ
к BaseSpace® – платформе для геномного анализа Illumina. Эта платформа предоставляет возможность выгрузки данных в реальном времени, простые инструменты для анализа данных, мониторинг цикла с подключением к сети Интернет и надежное решение для хранения с варьируемым размером. Пакет инструментов для анализа данных и растущий список приложений для BaseSpace от сторонних разработчиков дают исследователям возможность настраивать информационные системы индивидуально. Платформа BaseSpace обеспечивает также быстрый и простой обмен данными с коллегами или клиентами. Для получения более подробной информации посетите веб-сайт по адресу www.illumina.com/basespace. Малое время выполнения цикла, чтобы на получение результатов уходили не дни, а часы, система MiSeq в сочетании с быстрой подготовкой библиотеки с легкостью ускоряет время выполнения цикла. Подготовьте библиотеку секвенирования за 90 мин с помощью реактивов для подготовки Nextera®, а затем перейдите к автоматизированной клональной амплификации, секвенированию и распознаванию оснований, прошедших оценку качества, что на приборе MiSeq займет всего лишь 4 ч. Выравнивание последовательностей можно выполнить непосредственно на встроенном компьютере прибора с помощью программного обеспечения MiSeq Reporter или на платформе BaseSpace в течение 3 ч.
Революционный рабочий процесс системы MiSeq сокращает время выполнения цикла и позволяет проводить настольное секвенирование нового поколения. Образцы были подготовлены с помощью комплекта подготовки библиотеки Nextera XT. Секвенирование продолжительностью 4 ч включает в себя генерацию кластеров, секвенирование и распознавание оснований, прошедших оценку качества, со сканированием двух поверхностей для цикла с 1 × 36 пар оснований на системе MiSeq с программным обеспечением MiSeq Control v2.4.
Рабочий процесс включает следующие стадии:
- Подготовка библиотеки (добавление адаптеров для секвенирования
и подготовка библиотек к секвенированию). - Секвенирование (перенос библиотек в проточную кювету и секвенирование).
- Анализ данных (обработка данных/ добавление комментариев, составление отчета в приборе).
- Получение окончательных результатов (интерпретация и составление отчета о биологическом контексте).
Благодаря меньшему времени выполнения цикла и упрощенным рабочим процессам система MiSeq предлагает экономичную альтернативу капиллярному электрофорезу (КЭ) для таких областей применения, как целенаправленное ресеквенирование, проверка клонов и секвенирование ампликона (Nelson M. C., Morrison H. G., Benjamino J., SGrim. L., Graf J., 2014). Рабочие процессы анализа также оптимизированы для секвенирования генома малого размера, метагеномного анализа 16S рРНК, секвенирования РНК, секвенирования человеческого лейкоцитарного антигена (HLA), судебномедицинской экспертизы, преимплантационного генетического скрининга (ПГС) и преимплантационной генетической диагностики (ПГД), а также для областей применения с крайне сложной структурой, таких как TruSeq® Custom Amplicon и TruSeq Custom Enrichment (Frey K. G., Herrera-Galeano J. E., Redden C. L., Luu T. V., Servetas S. L., Mateczun A. J., Mokashi V. P., Bishop-Lilly K. A., 2014). Настраиваемая длина считывания, различные типы проточных кювет и возможность выбора считывания одиночных или парных концевых фрагментов обеспечивают беспрецедентную адаптируемость, благодаря чему выходные данные соответствуют широкому ряду потребностей, возникающих в ходе экспериментов.
1.3 БАКТЕРИОЦИНЫ ‒ БЕЛКИ, ПОДАВЛЯЮЩИЕ ЖИЗНЕДЕЯТЕЛЬНОСТЬ КЛЕТОК
ПАТОГЕННЫХ ШТАММОВ
В настоящее время антибиотикорезистентность является одним из основных глобальных вызовов, с которым столкнулось человечество. Трудность заключается в том, что в мире микроорганизмов все процессы изменчивости
и отбора происходят чаще и проявляются быстрее, а значит, вновь созданный препарат для борьбы с инфекцией необязательно будет абсолютной панацеей (Zhao W., Wang B., Yu G., 2018).
Антибиотикорезистентность обусловлена различными факторами: микроорганизмы научились либо изменять ту мишень, на которую направлено действие антибиотика, либо разрушать антибиотик, производя соответствующие ферменты. В то же время большинство бактериоцинов воздействуют в основном на клеточную стенку бактерий. Поскольку изменить ее основные характеристики и выработать устойчивость микроорганизму не так-то просто. Поэтому резистентность к антимикробным пептидам если и возникает, то в редких случаях и зачастую является временной. В этом и заключается преимущество подобных веществ перед традиционными антибиотиками (Vieco-Saiz N., Belguesmia Y., Raspoet R., Auclair E., Gancel F., Kempf I., Drider D., 2019).
Бактериоцины (антимикробные пептиды) в определенном смысле также являются антибиотиками, но иного происхождения. Это синтезируемые на рибосомах короткие последовательности аминокислот, которые в силу своих физико-химических свойств эффективно воздействуют на микроорганизмы, подавляя их развитие. Поскольку способностью синтезировать такие пептиды обладают многие живые существа, она рассматривается как наиболее древний и универсальный механизм защиты организма от чужеродных агентов.
Бактериоцины отличаются от классических антибиотиков тремя основными свойствами: синтез бактериоцинов происходит на рибосомах, бактериоцины обладают специфическим спектром действия, каждый бактериоцин имеет свой собственный специализированный иммунный белок.
Бактериоцины представляют собой комплекс пептидов с молекулярной массой от 2 до 35 кДа, существенно отличающихся друг от друга по физико-химическим характеристикам и биологическим эффектам (Sultimova T. D., Zakharov E. V., 2016). Выявлено, что на проявление антагонистической активности бактериоцинов, продуцируемых молочнокислыми бактериями, влияют температура, электрическое поле, рН, состав, консистенция среды, присутствие Ca2+ и Mg2+ и другие факторы (Kamarajan P., Hayami T., Matte B., Liu Y., Danciu T., Ramamoorthy A., Worden F., Kapila S., Kapila Y., Nisin Z. P., 2015; De Vuyst L., Leroy F., 2007). К числу общих свойств бактериоцинов относится их чувствительность к температуре, хотя это свойство также может варьировать в широких пределах. Некоторые разрушаются при температуре 48–50 С, другие кратковременно выдерживают температуру 60–70 С, а отдельные сохраняют активность даже при 100 С. Низин выдерживает кипячение до 120 С без потери активности. Бактериоцины чувствительны к воздействию протеаз и имеют различный молекулярный вес (30000–100000 Да). Белок бактериоцина связан с липополисахаридом клеточной оболочки, но только белковая часть молекулы обладает антибактериальной активностью (Oscariz J. C., Pissabarro A. G., 2011).
В настоящее время бактериоцины являются объектом изучения во многих странах. Исследования сосредоточены на свойствах новых бактериоцинов, выделенных у микроорганизмов разных таксономических групп (физико-химическая характеристика, спектр антибактериального действия, механизмы взаимодействия с микробной клеткой и макроорганизмом, возможности практического применения и т. д.). Как правило, изучение сопровождается определением молекулярно-генетических особенностей штаммов-продуцентов. На основании полученных результатов проводится классификация вещества (Kumariya R., Garsa A. K., Rajput Y. S., Sood S. K., Akhtar N., Patel S., 2011). Стоит отметить, что система международной классификации бактериоцинов несовершенна, ее продолжают уточнять и дополнять, так же, как и основные критерии принадлежности бактериальной субстанции к группе бактериоцинов. Особый интерес вызывают пептидсодержащие бактериоцины, как правило, имеющие широкий спектр действия и обладающие регуляторным влиянием на макроорганизм. Установлено, что как пробиотические штаммы, так и многие из изученных клинических культур обладают бактериоцинпродуцирующей активностью. Скрининг новых безопасных штаммов-продуцентов бактериоцинов, выявление их прикладного потенциала (пробиотики, анибактериальные средства, консерванты биопрепаратов, пищевых продуктов и др.) и получение соответствующих бактериоцинов является весьма актуальным и перспективным.
Использование бактериоцинов является одним из активно разрабатываемых альтернативных подходов к консервированию продуктов питания. Как известно, бактериоцины тесно ассоциированы с пищевыми продуктами
и имеют «GRAS» (Generally Recognized As Safe) статус, что определяет их как абсолютно безопасные для здоровья человека и животных (Taguchi S., Ooi T., Mizuno K., Matsusaki H., 2015; Vijayakumar P. P., Muriana P. M., 2015). Кроме того, бактериоцины в отличие от антибиотиков полностью расщепляются
в организме, и вероятность возникновения осложнений от бактериоцинов минимальна, а применение антибиотиков имеет для человека отрицательные последствия. Однако сферы применения известных форм бактериоцинов, как уже отмечалось, ограничены из-за низкой активности при нейтральных или щелочных значениях pH. Поэтому поиск бактериоцинов с улучшенными физико-химическими свойствами и широким антимикробным спектром представляет большой интерес не только для пищевой промышленности, но и для фармацевтической при создании альтернативных антибиотиков, что является одной из самых актуальных проблем в области микробиологии и биотехнологии.
1.3.1 Классификация бактериоцинов
Существует универсальная классификация бактериоцинов. Она включает четыре группы, различающиеся структурой, физико-химическими свойствами, особенностями синтеза и секреции (Rea M. C., Ross R. P., Cotter P. D., Hill C., 2011). Однако четвертый класс, в который входили бактериоцины в виде крупных комплексов с углеводными или липидными фрагментами, стали называть бактериолизинами (состоят из лейконоцина S и лактоцина 27) (Gulluce M., Karadayi M., Baris O., 2013), был расформирован. Таким образом, бактериоцины исследователи разделяют на три класса: I, II, III (таблица 4) (Liu W., Pang H., Zhang H., Cai Y., 2014).
Таблица 4 – Классификация бактериоцинов
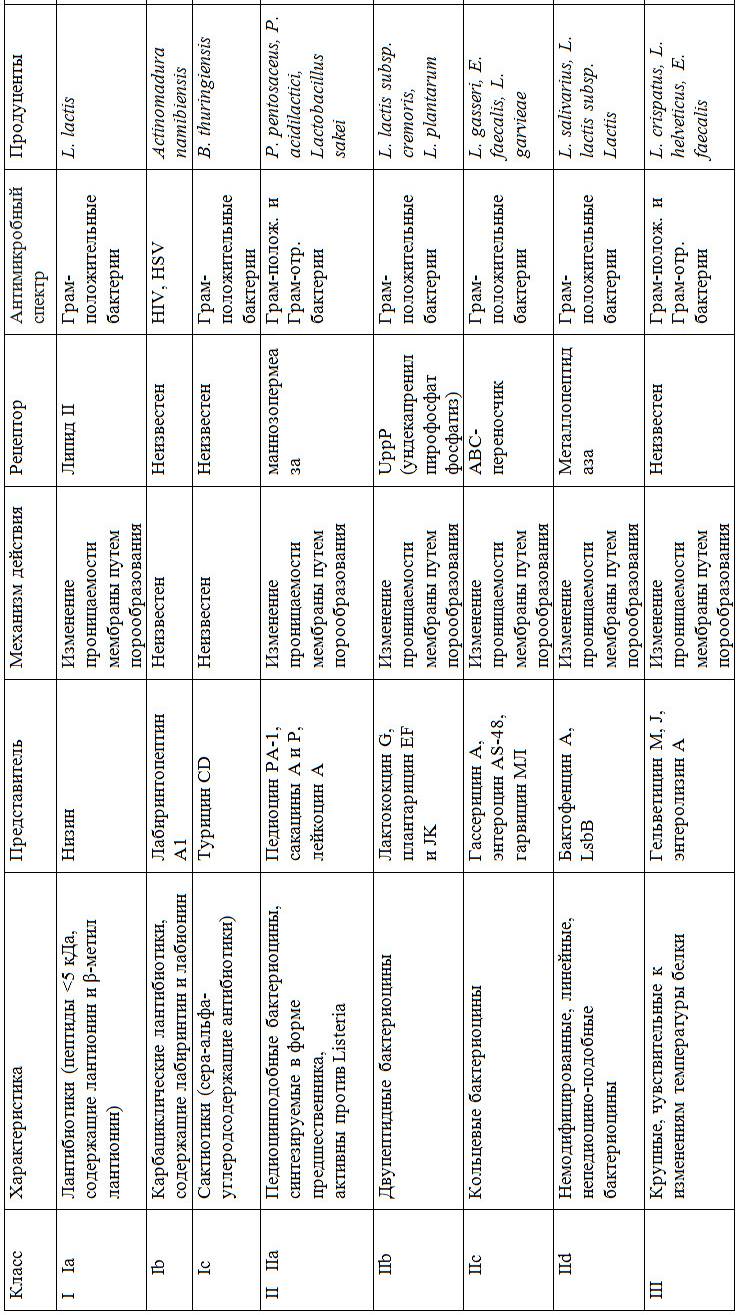
1.3.1.1 Бактериоцины I класса
Обычно состоят из 19–50 аминокислот и обычно сильно посттрансляционно модифицированы.
В результате образуются нетипичные аминокислоты, такие как лантионин, β-метиллантионин, дегидробутирин, дегидроаланин и лабиринтин (Cuozzo S., Sesma F., 2001; Parada J. L., Caron C. R., Medeiros A. B. P., Soccol C. R., 2007).
К этой группе относятся низины, образуемые Lactococcus (Streptococcus) lactis, эпидермин, галлидермин, продуцентом которого является St. gallinarum, а также субтилин (B. subtilis). Примером первого класса бактериоцинов является низин лантибиотик, секретируемый бактериями Lactococcus lactis. Содержит
34 остатка аминокислот (из них 30 остатков кислот – Lis, His, Asp, Ser, Pro, Gli, Ala, Val, Met, Ile, Leu), а также остатки редко встречающихся серосодержащих аминокислот: лантионина, -метиллантионина и ненасыщенных аминокислот дегидроаланина и -метилдегидроаланина. Низин обладает бактерицидным действием на грамположительные микроорганизмы: стрептококки различных серологических групп, стафилококки, пневмококки, микобактерии, споровые аэробные и анаэробные бактерии. Обладая способностью адсорбироваться на поверхности спор чувствительных к нему спорообразующих микроорганизмов, низин нарушает проницаемость спор, снижает устойчивость спор к воздействию высоких температур. Из аэробных споровых бактерий рода Bacillus к низину наиболее чувствительны B. stearothermophilus, B. coagulans, B. cereus, вызызвающие порчу молочных продуктов. Также чувствительны к действию низина споровые анаэробы рода Clostridium.
Класс I подразделяют на три подкласса:
Класс Ia, или лантибиотики, – низкомолекулярные катионные, гидрофобные, температуроустойчивые пептиды, в состав которых входят такие редкие тиоэфирные аминокислоты, как лантионин и -метиллантионин, дегидроаланин, дегидробутирин. Несколько тиоэфирных мостиков замещают дисульфидные связи и придают этим белковым соединениям некоторую полициклическую структуру. Низин является наиболее популярным бактериоцином класса Ia (Le Lay C., Dridi L., Bergeron M. G., Ouellette M., Fliss I., 2016).
Класс Ib – лабиринтопептины.
Класс Ic – сантибиотики (Cuozzo S., Sesma F., 2001).
1.3.1.2 Бактериоцины II класса
Большинство бактериоцинов относится ко II классу. Этот класс содержит небольшие термостабильные, немодифицированные, сохраняющие свою активность при широком диапазоне рН (3–9), слабо иммуногенные и нетоксичные для человека и животных (Dicks L. M. T., Balla E., Toit M., 2013; Oppegard, Rogne P., Emanuelsen L., Kristiansen P. E., Fimland G., Nissen-Meyer J., 2007). Бактериоцины II класса чаще всего синтезируются как препептиды, а затем без модификации их аминокислотных остатков, на стадии секреции из клетки продуцентов утрачивают свои лидирующие компоненты. Гены, необходимые для синтеза бактериоцинов II класса (регуляторные, гены, кодирующие препептид, обеспечивающие транспорт через мембрану и устойчивость бактерий к собственному антимикробному пептиду), располагаются в плазмидах или в хромосоме, а иногда в транспозоноподобных генетических модулях ДНК-содержащих бактериофагов (Farias R. N., Farias M. E., Holgado R., 2005). Бактериоцины II класса по химическому строению, способу выведения из клетки продуцента, спектру антимикробной активности разделяют на четыре подкласса: IIa, IIb, IIc, IId (Сультимова Т. Д. , 2006; Farias R. N., Farias M. E., Holgado R., 2005; Oscariz J. C., Pissabarro A. G., 2011).
Класс IIa – педиоцинподобные бактериоцины. Бактериоцины подкласса IIa наиболее хорошо изучены (Drider D., Fimland G., Hechard Y., McMullen L. M., Prevost H., 2006). Во время экспорта через клеточную мембрану от молекулы их препептидов отрезается N-терминальная лидирующая последовательность
с двумя глициновыми аминокислотными остатками на конце (Ермоленко Е. И., 2009). Педиоцинподобные бактериоцины имеют консенсусную последовательность YGNGV-C в N-конце.
После секреции из клетки эти бактериоцины представляют собой пептиды с молекулярной массой до 10 кДа, образующие, как правило, один дисульфидный мостик. После попадания в окружающую среду бактериоцины данного подкласса атакуют клетку-мишень за счет электростатического гидрофобного взаимодействия с последующим проникновением внутрь чувствительной клетки. Для специфического распознавания бактериоцином клетки-мишени необходимо присутствие на ее клеточной мембране маннозопротеиновых рецепторов, являющихся частью фосфотрансферазной системы. При помощи C-части пептиды проникают в мембрану и действуют на клетку-мишень, что определяет антибактериальную активность и специфичность бактериоцина. Показано, что степень проникновения зависит от заряда липосом, а также от расположения аминокислотных остатков в С-части молекулы бактериоцина. Бактериоцины IIa типа формируют ионоселективные поры в клетках-мишенях, вызывая выход из последних ионов калия, аминокислот и других низкомолекулярных веществ. Это приводит к изменению ионного баланса, уровня рН в клетке и последующей быстрой потере внутриклеточной АТФ (Ермоленко Е. И., 2009).
Класс IIb – двупептидные бактериоцины, состоят из двух различных субъединиц или пептидов, действующих вместе или раздельно как синергисты (Farias R. N., Farias M. E., Holgado R., 2005; Kraus A., Peschel D., 2006; Oscariz J. C., Pissabarro A. G., 2011). До настоящего времени, несмотря на способность проявлять антимикробную активность самостоятельно и расположение
в различных генах, эти вещества не были выделены.
Класс IIc – кольцевые бактериоцины. Примером является бактериоцин AS-48. Он является циклическим пептидом с молекулярной массой 7,2 кДа, представляет собой полипептидную цепь из 70 аминокислотных остатков, замкнутых в кольцо. Известно, что подобная молекула обладает высокой конформационной изменчивостью (Farias R. N., Farias M. E., Holgado R., 2005). Это свойство позволяет молекуле бактериоцина AS-48 связываться с поверхностными структурами широкого спектра бактерий.
Класс IId – линейные, не относящиеся к педиоцино-подобным бактериоцинам (Belguesmia Y., Naghmouchi K., Chihib N.-E., Drider D., 2011; Drider D., Fimland G., Hechard Y., McMullen L. M., Prevost H., 2006; Oppegard C., Rogne P., Emanuelsen L., Kristiansen P. E., Fimland G., NissenMeyer J., 2007). Бактофенцин А является новым бактериоцином класса IId, обладает определенными уникальными функциями. Это положительно заряженный антимикробный пептид, имеет сходство с некоторыми эукариотическими катионными антимикробными пептидами (O’Connor P. M., O’Shea E. F., Cotter P. D., Hill C., Ross R. P., 2018; O’Shea E. F., O’Connor P. M., O’Sullivan O., Cotter P. D., Ross R. P., Hill C., 2013).
1.3.1.3 Бактериоцины III класса
Бактериоцины этого класса представляют собой крупные чувствительные к изменениям температуры белки с молекулярной массой более 30 кДа. Они способные разрушать муреин клеточной стенки. Колицин является одним из примеров бактериоцинов класса III, продуцируемых E.coli. К классу III бактериоцинов также относятся гельветицин М, гельветицин J и энтеролизин А, продуцируемые Lactobacillus crispatus, Lactobacillus helveticus и E. faecalis, соответственно (Nilsen T., Nes I. F., Holo H., 2003; Sun Z., Wang X., Zhang X., Wu H., Zou Y., Li P., Sun C., Xu W., Liu F., Wang D., 2018; Thompson J. K., Collins M. A., Mercer W. D., 1996). Гельветицин М эффективен против грамположительных
и грамотрицательных бактерий (Sun Z., Wang X., Zhang X., Wu H., Zou Y., Li P., Sun C., Xu W., Liu F., Wang D., 2018).
Классификация бактериоцинов дает возможность систематизировать информацию о природе и механизмах действия соответствующих групп антимикробных пептидов. Это позволяет осуществлять подбор комбинаций антимикробных пептидов для лечения и профилактики различной инфекционной патологии (Ankaiah D., Palanichamy E., Antonyraj C. B., Ayyanna R., Perumal V., Ahamed S. B., Arul V., 2018).
1.3.2 Механизм действия бактериоцинов
Учитывая большое разнообразие химического строения бактериоцинов, можно предположить, что они воздействуют на жизненно важные функции чувствительных клеток, но большинство действуют, образуя в мембране поры или каналы, способствующие нарушению мембранного потенциала чувствительных клеток (Сультимова Т. Д., 2006; Surugau L. N., 2011). Анионные липиды цитоплазматической мембраны являются основными рецепторами бактериоцинов молочнокислых бактерий для стимуляции спорообразования.
Бактериоцины I класса могут вызывать формирование пор по типу клиноподобной модели, в то время как бактериоцины II класса способны функционировать, создавая поры, посредством которых пептиды располагаются параллельно мембранной поверхности и оказывают влияние на структуру мембран (Chatterjee M., Raichaudhuri A., 2017; Lopez-Lara I. M., Geiger O., 2017). Наиболее хорошо изучен механизм действия бактериоцинов на примере низина (рисунок 2).
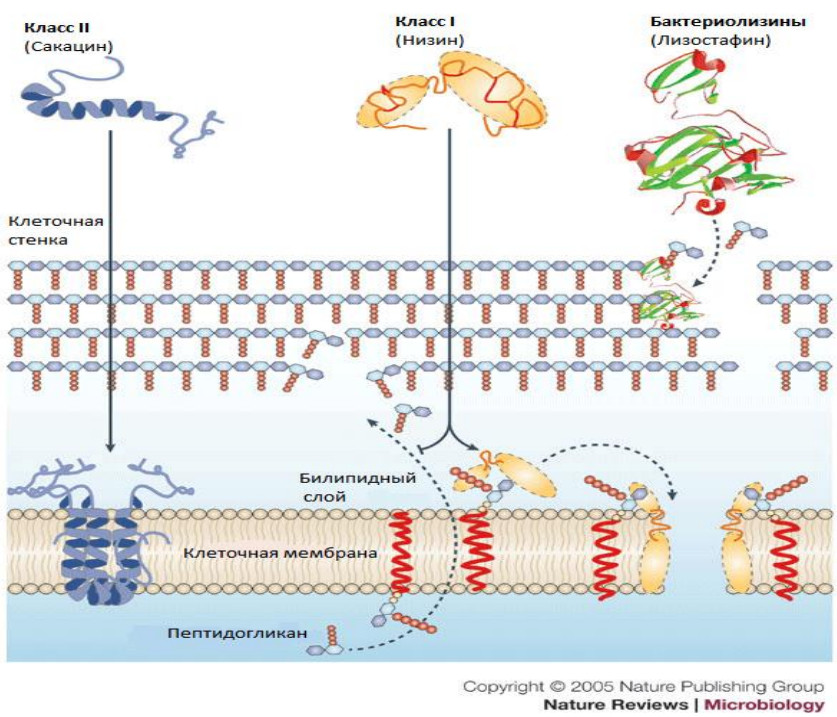
Рисунок 2 – Механизм действия бактериоцинов
При изучении действия низина стало ясно, что часть C-конца молекулы деполяризует цитоплазматическую мембрану, что ведет к мгновенной остановке синтеза компонентов клеточной стенки и всех биосинтетических процессов
в клетке. Основной характер действия низина это взаимодействие с отрицательно заряженными мембранами (билипидный слой), предпочтительно с мембранами, содержащими анионные липиды. Грамположительные бактерии имеют более высокие концентрации анионного липида в их цитоплазматической мембране по сравнению с грамотрицательными. Бактериоцин сначала связывает его C-конец с анионными липидами, после чего N-конец опускается в липид мембраны и пептид принимает параллельное положение на мембранной поверхности. И, в конечном счете, целый пептид имеет возможность переместиться по всей длине поры (Surugau L. N., 2011). Низин ингибирует синтез муреина клеточной стенки. В результате действия антибиотика происходит накопление промежуточного липида, с которым низин образует прочный комплекс, что и приводит к остановке дальнейшего синтеза пептидогликана бактериальной стенки. Низин может вызывать лизис клеток вследствие необратимой деструкции клеточной мембраны, которая регулирует осмотическое давление клетки. Он изменяет поверхностное натяжение цитоплазматической мембраны, что приводит к нарушению клеточной проницаемости и, как следствие этого,
к бактериальному лизису.
Низин ингибирует синтез муреина клеточной стенки. В результате действия антибиотика происходит накопление промежуточного липида, с которым низин образует прочный комплекс, что и приводит к остановке дальнейшего синтеза пептидогликана бактериальной стенки. Низин может вызывать лизис клеток вследствие необратимой деструкции клеточной мембраны, которая регулирует осмотическое давление клетки. Он изменяет поверхностное натяжение цитоплазматической мембраны, что приводит к нарушению клеточной проницаемости и, как следствие этого, к бактериальному лизису.
1.3.3 Бактериальные штаммы – продуценты бактериоцинов
Известно, что многие микроорганизмы способны к синтезу бактериоцинов. Но наибольший интерес с точки зрения применения в качестве новых консервантов и антибактериальных препаратов представляют бактериоцины молочнокислых бактерий. Исследования в области изучения бакте риоцинов у молочнокислых бактерий начались в 1930-х гг. прошлого столетия с работы по низину, продуцентом которого были лактококки L. lactis subsp. lactis (Yanagida F., Chen Y., Shinohara T. J., 2005). К настоящему времени изучено и полностью охарактеризовано большое количество бактериоцинов, синтезируемых как грамположительными, так и грамотрицательными бактериями. Продуцирующие бактериоцин молочнокислые бактерии включают представителей разных родов: Lactococcus, Lactobacillus, Leuconostoc, Pediococcus и Streptococcus. Синтез бактериоцинов штаммоспецифичен. Наибольшее количество бактериоцинобразующих бактерий было выделено из молочных и мясных ферментированных продуктов, силоса, но ряд штаммов (Enterococcus durans, Lactobacillus animalis, Leuconostoc sp.) были выделены из почвенных образцов (Beasley S., Saris P., 2004). Имеются сведения, что штаммы L. lactis subsp. lactis, продуцирующие низин, были выделены из женского молока (Fimland G., Eijsink, Nissen’Meyer J., 2002).
Микроорганизмы – продуценты бактериоцинов – имеют систему защиты от своих собственных продуктов с помощью экспрессии генов иммунных белков. Чувствительность продуцента варьирует и зависит от уровня продукции бактериоцина (Willey M., van der Donk W., 2007). Иммунные белки проявляют значительную вариабельность: у практически одинаковых бактериоцинов могут быть абсолютно разные иммунные белки. Для низина известен NisI – белок
с липопротеиновой сигнальной последовательностью, кодируемый геном nisI
в опероне биосинтеза низина. Белок NisI прикрепляется к внешней поверхности мембраны клетки-продуцента, где связывает низин, уменьшая его локальную концентрацию (Mapelli C., Musatti A., Barbiroli A., Saini S., Bras J., Cavicchioli D., Rollini M., 2019).
В таблице 5 перечислены представители наиболее хорошо изученного
и широко представленного класса II бактериоцинов и их продуцентов.
Скрининг бактериоцин-продуцирующих микроорганизмов в природных объектах является самым часто используемым методом получения новых продуцентов бактериоцинов. Молочнокислые бактерии широко распространены
в природе: их можно обнаружить в почве, на разлагающихся остатках животного и растительного происхождения, в кишечнике позвоночных, в молоке и молочных продуктах. Вместе с растениями и пищей они попадают в желудочно-кишечные тракты человека и животных, составляя его микробиоту. В зависимости от источников разработаны и оптимизированы для использования различные системы скрининга бактериоцинов.
Таблица 5 – Представители класса II бактериоцинов и их продуцентов
| Бактериоцины Класса IIa | ||
| Бактериоцин | Продуцент | Ссылка |
| Сакацин A
Сакацин 674 Лактицин Q Лактококцин А Лактококцин 972 Лактококцин DR Карнобактериоцин А Карнобактериоцин В2 Курвацин А Диверцин V41 Дивергицин Лактококцин ММFII Лейкоцин А Мезентерицин Y105 Плантарицин 423 Педиоцин РА1 Педиоцин АсН Сакацин G |
Lactobacillus sakei Lb 706
L. sake Lb764 Lactococcus lactis QU 5 L. lactis subsp. cremoris LMG 2130 L. lactis subsp. lactis IPLA 972 L. lactis subsp. lactis ADRIA 85L030 Carnobacterium piscicola LV17A C. piscicola LV17 Lactobacillus curvatus LTH1174 Carnobacterium divergens М35 C. divergens M35 L. lactis MMFII Leuconostoc gelidum UAL 187 L. mesenteroides Y105 Lactobacillus plantarum 423 Pediococcus acidilactici PAC 1.0 P. acidilactici AcH Lactobacillus sake 2512 |
[134, 135]
[136] [137] [138] [139] [140] [141] [142] [143] [144] [145] [146] [147] [148] [149] [150] [151] [152] |
| Бактериоцины Класса IIb | ||
| Лактококцин G
Лактококцин MN Лактококцин Q Плантарицин E/F Плантарицин J/K Плантарицин NC8 Лактацин F Лактоцин 705 Термофилин 13 Лейкоцин Н |
L. lactis LMGT2081
L. lactis 9B4 L. lactis QU 4 Lactobacillus plantarum C11 L. plantarum C11 L. plantarum NC8 Lactobacillus johnsonii VPI11088 Lactobacillus casei CRL 705 Streptococcus thermophilus Sfi13 Leuconostoc MF215B |
[153]
[153] [154] [155] [156] [157] [158] [159] [160] [161] |
Приведем несколько примеров выделения продуцентов бактериоцинов из различных источников. Штамм Leuconostoc gelidiumstain UAL187 был выделен из упакованного мяса. Он синтезирует бактериоцин лейкоцин А, который активен против молочнокислых бактерий, Listeria и E. faecalis. Кодирующую последовательность лейкоцина А находится в плазмиде pLG7.6. Культуру растили,
и лейкоцин выделяли из супернатанта культуры, получив 2,06 мг белка, и этот бактериоцин состоял из 37 аминокислот. Теоретическая молекулярная масса, рассчитанная по предполагаемой аминокислотной последовательности, составила 3932,3 Да, что на 2 Да больше, чем молекулярная масса, определенная из масс-спектрометрического анализа выделенного лейкоцина А. Уменьшение молекулярной массы объясняется потерей двух протонов при образовании дисульфидной связи между двумя остатками цистеина во время созревания белка (US20100129864 A1, 2010; US6403082 B1, 2002).
Штамм Brochothrix campestris ATCC 43754 был выделен из образца почвы. Этот штамм продуцирует бактериоцин брохоцин-С.
Последовательность, кодирующая брохоцин, была выделена из хромосомной ДНК. Бактериоцин проявлял активность в отношении возбудителя ботулизма Clostridium botulinum и широкого спектра грамположительных бактерий. Брохоцин-С был стабилен при 121 °С в течение 15 мин и инактивировался ферментативной обработкой (US20100129864 A1, 2010; US6403082 B1, 2002; Siragusa G. R., Cutter C. N., 1993).
E. faecium 900 продуцирует энтероцин 900. Ген экспрессии белка находится на его хромосоме. Энтероцин состоит из 71 аминокислотных остатков и проявляет активность против широкого спектра грамположительных и грамотрицательных бактерий (US20100129864 A1, 2010; US6403082 B1, 2002; Franz C. M., Schillinger U., Holzapfel W. H., 1996).
Streptomyces scopuliridis RB72 был выделен из почвы. Это грамположительная неспоровая неподвижная аэробная бактерия, продуцирующая бактерицидно-подобное вещество широкого спектра действия (BLIS RB72), которое убивает/ингибирует грамположительные и грамотрицательные бактерии. BLIS RB72 имеет молекулярную массу 27 кДа и демонстрирует остаточную активность при температуре от минус 20 до 60 °C и рН 3–9 и активен против бактерий при рН 7 (US20100286030 A1, 2010; Farris M. H., Duffy C., Findlay R. H., Olson J. B., 2011).
Bifidobacterium longum DJO10A – это бактериальный штамм с пробиотическими свойствами, выделенный из человека. Эта бактерия продуцирует лантибиотик, который активен против грамположительных и грамотрицательных бактерий. Неочищенный лантибиотик, выделенный из культуры, способен выдержать при 100 °C в течение 10 мин и обработку трипсином – при 25 °C в течение 24 ч (US7960505 B2, 2011).
Lactobacillus casei MRTL3 выделен из образцов сырого молока молочных ферм г. Харьяна, Индия. Оптимальная температура для культурального роста 38 °С. Бактериальный штамм Lactobacillus casei MRTL3 продуцирует бактериоцин, проявляющий стабильность при 100 °C в течение 5 мин, и активен
в отношении грамположительных бактерий (Staphylococcus aureus ATCC 6538P, S. Epidermidis ATCC 12228, Bacillus cereus ATCC 11770, L. monocytogenes MTCC 657 и L. innocua ATCC 33090) и грамотрицательных бактерий (Shigella flexneri ATCC 9199, Salmonella typhi MTCC 733 и Pseudomonas aeruginosa ATCC 15442). Наряду с бактериоцином лактобациллы продуцирует гликолипиды, обладающие антимикробными свойствами (Sharma D., Singh Saharan B.). Штамм Bacillus coagulans был выделен из ризосферы. Максимальный рост этой бактерии был получен в среде MRS наряду с высокой продукцией бактериоцина. Очищенный бактериоцин имеет молекулярную массу 8 кДа. Концентрированный и лиофилизированный бактериоцин показал высокую антагонистическую активность против S. aureus, P. aeruginosa, E.coli и низкую активность в отношении S. typhi, L. monocytogenes и C. perfringins (Ranganath B. S., Sharmila T., Balasubramanyam B. V., 2015). Несмотря на то, что многие бактериоцин-продуцирующие бактерии были выделены из огромного разнообразия источников, ни один из них не был подготовлен для промышленного производства и не получил разрешения FDA для использования в пищевом производстве. Вновь выделенные бактериоцины-продуценты бактерий перечислены
в таблице 6.
Таблица 6 – Штаммы-продуценты бактериоцинов
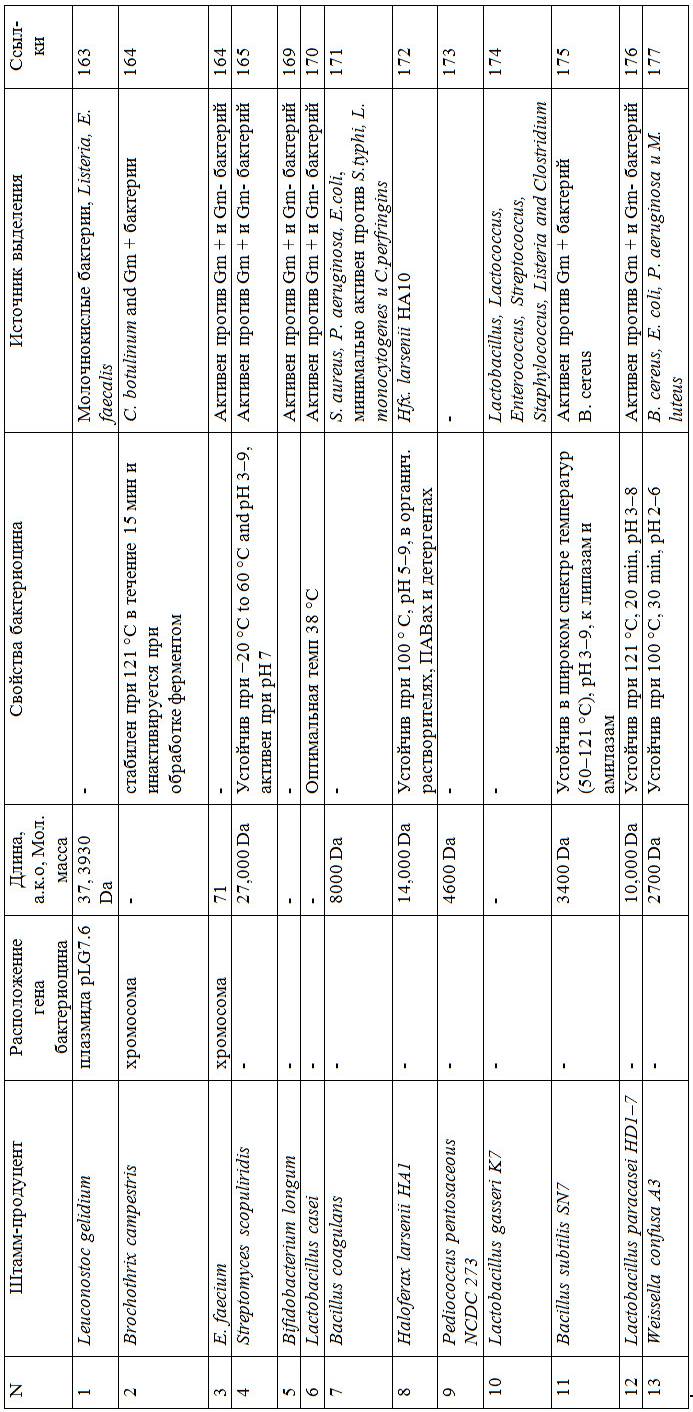
1.3.4 Рекомбинантные бактериоцины
Принимая во внимание успешные примеры производства рекомбинантных белков с использованием бактерий и дрожжевых клеток, также изучено производство рекомбинантных бактериоцинов. Как правило, гетерологические системы экспрессии бактериоцинов ограничены бактериями и клетками дрожжей.
1.3.4.1 Экспрессия в бактериальных клетках.
Гетерологическая экспрессия трех различных бактериоцинов класса II (Sakacin P, Pediocin PA-1 и Piscicolin 61) была успешно проведена в L. sake Lb790 (pSAK20). Этот штамм-хозяин не имеет каких-либо нативных генов продукции бактериоцинов. Две плазмиды – pSAK20, содержащая ген бактериоцина для гетерологичной экспрессии, и производная плазмиды pLPV111, содержащая структурные гены и гены иммунитета для представляющего интерес бактериоцина, – были совместно экспрессированы. С использованием этой системы различные бактериоцины продуцировались на уровнях, равных или более высоких, чем уровни, полученные с соответствующими штаммами-продуцентами дикого типа. Антимикробную активность количественно выражали в арбитражных единицах (АU; из общепринятого англоязычного обозначения arbitration units, аналогичного аббревиатуре МПК – минимальная подавляющяя концентрация), отнесенных к 1 мл или 1 мг пробы. Максимальная активность бактериоцина, полученная в жидкой культуре, для Sakacin P, составила 24, Pediocin PA-1 – 32, а Piscicolin 61 – 3,9 kAU (1 кAU мл–1 = 1000 AU/мл), что было выше, чем у соответствующего штамма дикого типа (Axelsson L., Katla T., Bjørnslett M., Eijsink V. G., Holck A., 1998). Бактериоцин энтероцин A (Ent A)
и его иммунный белок Ent I (EntIA) из E. faecium T136 были клонированы для коэкспрессии сигнального пептида белка Usp45 в L. lactis MG1363. Три плазмиды pSIP409UAI, pSIP411UAI (индуцибельная) и pMGUAI (конститутивная) были сконструированы и трансформированы в L. sakei Lb790, L. plantarum NC8 и L. casei CECT475. Определяли экспрессию бактериоцина в супернатантах культуры (мкг/мг сухого веса клеток), антимикробную активность (BU/мг сухого веса клеток) и специфическую антимикробную активность (BU/мкг EntA). Было отмечено, что эти значения варьировались для всех трех штаммов-хозяинов и были выше в L. casei CECT475, который можно рассматривать как фабрику перспективных бактериальных клеток для гетерологичной экспрессии бактериоцина (Jiménez J. J., Diep D. B., Borrero J., Gútiez L., Arbulu S., Nes I. F., Herranz C., Cintas L. M., Hernández P. E., 2015). L. mesentemides Y105 продуцируют антилистериальный бактериоцин мезентерицин Y105. В нативном штамме бактериоцин экспрессировался в секреторном режиме с использованием специальной транспортной системы (DTS от dedicated transport system). Гетерологичная экспрессия мезентерицина Y105 была проведена в E. coli DH5α,
L. mesentroides DSM 20484 и L. lactis штаммов IL1403 и RD230 как для DTS, так и для общего пути секреции (GSP от general secretion pathway). Гетерологическая экспрессия в L. lactis не удалась. В E.coli рекомбинантный белок был токсичным для клеток-хозяев, что приводило к лизису клеток. Однако секретируемый белок проявлял умеренную антагонистическую активность против Listeria. Плазмидная конструкция, содержащая как ген мезентерицина, так и его иммунный ген, а также три предполагаемых гена мезентрицина, которые участвуют
в синтезе транспортной системы DTS, как было показано, экспрессирует рекомбинантный белок (Biet F., Berjeaud J. M., Worobo R. W., Cenatiempo Y., Fremaux C., 1998).
L. plantarum ZJ316 экспрессирует бактериоцин плантарицин NC8. Он состоит из двух пептидов – PLNC8α и PLNC8β, гетерологически экспрессированных в E. coli BL21 (DE3). Рекомбинантные пептиды PLNC8α и PLNC8β экспрессировали в виде слитых белков His6-tag для легкой очистки с помощью аффинной хроматографии на колонке Ni2+. Для получения нативных N-концевых рекомбинантных пептидов His-меченые белки расщепляли энтерокиназой и очищали с использованием Ni-NTA Sefinose™ ResinKit. По результатам электрофоретического анализа молекулярные массы рекомбинантных пептидов составляли 3,58 и 4 кДа, соответственно. Выходы чистых белков составили
2–2,5 и 1,5–2 мг/л соответственно для PLNC8α и PLNC8β. Очищенные белки показали бактерицидную активность против возбудителя Salmonella spp. (Jiang H., Li P., Gu Q., 2016).
Geobacillus thermodenitrificans NG-80-2 был выделен из образца глубинной поверхности нефтяного месторождения Даганг в Китае. Температура недр была около 50–60 °С. Как было показано, в образце обнаружены бактериоцины широкого спектра геобациллин I и II. Кодирующие последовательности геобациллина I и II были амплифицированы и гетерологически экспрессированы в клетки E. coli BL21 (DE3). Его меченые рекомбинантные белки были очищены, а их структуры определены. Геобациллин I проявлял активность против многих антибиотикорезистентных патогенных микроорганизмов в отличие от геобациллина II. Геобациллин I ингибирует бактерии подобно низину, а именно, он ингибирует синтез пептидогликана путем связывания с молекулой липида II в зависимости от дозы (US9326523 B2, 2016).
1.3.4.2 Экспрессия в дрожжевых клетках
L. plantarum 423 содержит оперон, экспрессирующий бактериоцин. Этот оперон имеет четыре гена, первая открытая рамка считывания (ORF) включает plaA CDS, экспрессирующий плантарин 423. Вторая ORF имеет plaBen-кодирующий предполагаемый иммунный белок, PlaC и PlaD гены, идентичные оперону PA-1 педиоцина (PedC and PedD) и оперон, идентичный коагулину (CoaC и CaencoA). plaA кодирует пептид-предшественник длиною в 56 аминокислотных остатков, который состоит из зрелого белка (37 а.к.о.) и NH2-концевого лидерного пептида (19 а.к.о.). PlaA CDS был клонирован в челночный вектор для секреторной экспрессии (YEpPla423) в S. cerevisiae L5366 h. Секреторная экспрессия плантарицина 423 была подтверждена образованием зоны ингибирования над индикаторными пластинками L. monocytogenes LM1 (Van Reenen C. A., Chikindas M. L., Van Zyl W. H., Dicks L. M., 2003). Последовательность SEC-зависимого бактериоцина хирацина JM79 из Enterococcus hirae DCH5 была клонирована и экспрессирована в плазмиду PICZαA под промотором AOX1 и трансформирована в дрожжевой штамм Pichia pastoris X-33. Рекомбинантный дрожжевой клон X-33TH, содержащий плазмиду, секретировал биологически активный бактериоцин, как в минимальной среде с буферизованным метанолом (BMM), так и в среде с буферизованным комплексом метанола (BMMY). Продукция белка достигала своего максимума через 6 ч, концентрация составляла 6,57 мкг/мг CDW и 2,75 мкг/мг CDW в BMM и BMMY, соответственно (Sánchez J., Borrero J., Gómez-Sala B., Basanta A., Herranz C., Cintas L. M., Hernández P. E., 2008).
Ген энтероцина A (EntA) из E. faecium T136 клонировали в разные плазмиды дрожжей для секреторной экспрессии в P. pastoris, Kluyveromyces lactis, Hansenula polymorpha и Arxula adeninivorans. Самая высокая экспрессия энтероцина А достигается в P. pastoris X-33EA, K. lactis GG799EA и H. polymorpha KL8-1EA, что соответственно в 30, 7 и 3,2 раз выше, чем у нативного штамма. В аналогичных линиях антимикробная активность супернатантов из C. pastoris and K. lactis была в 432 и 59 раз выше, чем у нативного штамма. Антимикробная активность H. polymorpha и A. adeninivorans составила, соответственно, 85 и 19 % от активности нативного штамма. Специфическая антимикробная активность С. pastoris и K. LACTIS были в 14,4 и 8,4 раза выше, чем у E. faecium (Borrero J., Kunze G., Jiménez J. J., Böer E., Gútiez L., Herranz C., Cintas L. M., Hernández P. E., 2012).
Белки бактериоцина широкого спектра действия из различных бактерий SRCAM602, OR-7, E-760 и L-1077 были отобраны для секреторной экспрессии в P. pastoris. Оптимизированные по кодонам синтетические гены были клонированы в pPICZαA для секреторной экспрессии in P. pastoris X-33. Рекомбинантные белки из супернатанта не проявляли какой-либо антимикробной активности. Очищенные белки проявили свою антимикробную активность только после гидрофобной хроматографии. Масс-спектрометрический анализ очищенного белка показал более высокую молекулярную массу, чем ожидалось, из-за присутствия дополнительных посттрансляционных модификаций нативных дрожжей (Arbulu S., Jiménez J. J., Gútiez L., Cintas L. M., Herranz C., Hernández P. E., 2015).
Ген PedA, продуцирующий педиоцин, был клонирован в эписомальной плазмиды под сигнальную последовательность α-фактора для секреторной экспрессии в S. сerevisiae Y294. Рекомбинантный белок присутствовал в супернатанте культуры с молекулярной массой 4,6 кДа. Было обнаружено, что уровень гетерологичной экспрессии бактериоцина является низким, но он проявляет значительную антимикробную активность на L. monocytogenes B73. Рекомбинантный штамм дрожжей одновременно производил спирт во время брожения пива и вина, но также выделял бактериоцины. Эти секретированные бактериоцины могут ингибировать рост бактерий, вызывающих порчу пива и вина (Schoeman H., Vivier M. A., Dutoit M., Dicks L. M. T., Pretorius I. S., 1999).
Ген PedA, экспрессирующий бактериоцин педиоцин PА-1 из P. acidilactici, клонировали в плазмиду pPICZαA для секреторной экспрессии in P. pastoris штаммов X-33, GS115 и KM71H. Ферментация при встряхивании в шейере
в рекомбинантных штаммах позволяла получить белков концентрацией
74 мкг/мл без антимикробной активности. Вестерн-блот анализ показал наличие трех разных изоформ белков с молекулярной массой 7–8, 11 и 20 кДа, которые выше, чем у природного педиоцина PA-1 (4,6 кДа). Анализ рекомбинантного белка путем изоэлектрического фокусирования, секвенирования белка
и расщепления коллагеназой показал, что белок ведет себя подобно коллагену
и образует четные волокна. Причину такого специфического поведения рекомбинантного педиоцина еще предстоит выяснить (Beaulieu L., Groleau D., Miguez C. B., Jette J. F., Aomari H., Subirade M., 1999).
Продукция рекомбинантных бактериоцинов методом гетерологической экспрессии является относительно низкой и иногда приводит к низкой или даже к отсутствию антимикробной активности. Удивительно, что в хорошо зарекомендовавших себя платформах экспрессии дрожжевого белка, таких как P. pastoris, экспрессия рекомбинантного белка была очень низкой или бесполезно продуцировала инактивированные белки. Необходимо разрабатывать более сложные стратегии для повышения уровня экспрессии и антимикробной активности.
Промышленное применение бактериоцинов
Применение бактериоцинов преимущественно сосредоточено в пищевой промышленности. В связи с увеличением количества вновь открытых бактериоцинов, спектр их применения может быть расширен и в таких областях, как садоводство, биомедицина и личная гигиена. Последние 10-летние отчеты
о применении бактериоцина в различных областях кратко изложены ниже.
Например, штамм L. plantarum subsp. plantarum Zhang-LL (CGMCC
№ 6936) был выделен из бекона (г. Фуцзянь, Китай). Эта бактерия продуцирует бактериоцин, который может подавлять рост L. monocytogenes, проявляет высокую термостабильность, а также стабильность в кислотных и основных pH. И в то же время расщепляется пищеварительным соком и ферментами в желудке человека, что делает его идеальным пищевым биоконсервантом. Композиция, содержащая и бактериоцин из штамма Zhang-LL и низин, показала синергизм действия этих веществ (Hongxing L., Astrøm A. N., List T., Nilsson I. M., Johansson A., 2016; Zhang, Hongxing, Liu, Hui, Bai, Yongqiang, Xu, Hongyuan, Duan, Huixia, Hao, Yanfang, Xie, Yuanhong, Jin, Junhua, Xiong, Lixia, Gao, Xiuzhi, 2015).
Кукурузная мезга используется для производства биоэтанола. Такие лактобактерии, как Lactobacillus, Lactococcus, Weissella, Leuconostoc, Pediococcus
и Enterococcus бактерии, загрязняют мезгу, способствуя преобразованию крахмала в молочную и укусную кислоты, тем самым снижают продуктивность процесса и выход биоэтанола. Не молочнокислые бактерии, такие как Streptococcus, Staphylococcus, Bacillus и Clostridium являются общезагрязняющими микроорганизмами, которые влияют на общую эффективность ферментации. Для преодоления проблемы загрязнения бактериями, из кукурузной мезги был выделен бактериоцин-продуцирующий штамм L. plantarum GP15. Он производит бактериоцин GP15-cin широкого спектра действия. Использование GP15-cin уменьшило общее микробное загрязнение кукурузной мезги на 99,9999 %
(> 800 000 раз) в течение 2 ч, что значительно снижало уровень производства молочной кислоты и уксусной кислоты в процессе ферментации этанола.
LferInf – бактериофаг широкого спектра действия, обладающий антагонистической способностью против как кисломолочных, так и не кисломолочных бактерий. Состав, содержащий 1 % бактериофага LferInf и GP15-cin, при различных концентрациях патогенов способствовал снижению роста заражения бактериями до 0 в течение 25 мин, повышая при этом эффективность биоэтанольного брожения (US20140148379 A1, 2014; US20150320829 A1, 2015).
В рецептуре хлеба при добавлении 20 % штамма P. pentosaceus KTU05-9
к закваске, данный штамм Pediococcus продуцирует органические кислоты
и бактериоцины, которые ингибируют рост B. subtilis, придают вязкости хлебу и продлевают срок годности хлеба до 6 дней. Микробиотическая смесь, содержащая P. acidilactici KTU05-7, P. pentosaceus KTU05-8 и KTU05–10, распыленная на поверхности упакованного хлеба, ингибирует рост грибков и сохраняет его свежим в течение 8 дней (Cizeikiene D., Juodeikeine G., Paskevicius A., Bartkiene E., 2013).
Бактериоцины являются промышленно привлекательными веществами из-за их большого потенциала в качестве натуральных консервантов в пищевой, кормовой и косметической промышленностях. До сих пор низин является единственным бактериоцином, который был одобрен для применений в пищевой промышленности. Низин преимущественно используется в консервировании мяса и мясопродуктов, морепродуктов и овощей. Рынок низина в Азиатско-Тихоокеанском регионе быстро растет, за ним следует рынок Европы и Северной Америки. Ожидается, что к 2020 г. мировой рынок низина достигнет
545,5 млн долл., годовой темп роста (CAGR) в период 2015–2020 гг. составил 4,3 % (http://www.marketsandmarkets.com/PressReleases/nisin.asp).
Учитывая растущий спрос и низкую производительность низина (до
9960 IU/mL), необходимо применить меры для увеличения эффективности производства низина и снижения его себестоимости.
Для производства низина используют негенномодифицированные лактобактерии, с помощью мутагенеза целых клеток возможно получение высокопроизводительных штаммов. Разнообразные физические мутагены (γ-лучи
и излучение плазмы), химические мутагены (этилметилсульфат, алкилирующие агенты) и адаптационная эволюция может быть использована для создания негенномодифицированные лактобактерии (Juturu V., Wu J. C., 2018).
1.3.5 Бактериоцины, имеющие статус в международной системе безопасности микроорганизмов
Несмотря на значительное количество недавно открытых бактериоцин-продуцирующих бактерий широкого спектра действия, получить утверждение от правительственных агентств для их промышленного применение не удается. Учитывая открытие новых бактериоцинов с новыми свойствами, следует исследовать их применение в таких областях, как корма для животных, органические удобрения и охрана окружающей среды, а также в продуктах личной гигиены.
Бактериоцин низин ‒ единственный из бактериоцинов, с 1998 г. имеющий cтатус GRAS (Generally Recognized As Safe) и разрешенный для использования в качестве пищевой добавки (код Е234) (European Parliament. EC 248/97, 1997).
В результате широкого использования молочнокислых бактерий в традиционных ферментированных продуктах большая их часть считается безопасной согласно международной системе безопасности микроорганизмов (Generally Recognized As Safe, GRAS), предоставленной Американским агентством по контролю за продуктами и лекарствами (FDA). Европейский орган по безопасности пищевых продуктов (EFSA) также предоставил статус «Квалифицированная презумпция безопасности» (QPS) большинству родов молочнокислых бактерий, таким как Lactococcus, Lactobacillus, Leuconostoc, Pediococcus и некоторые виды Streptococcus (Question № EFSA-Q-2005-293) // EFSA J., 2007). Тем не менее, виды рода Enterococcus и некоторые виды Streptococcus являются патогенными, поэтому они не имеют статуса GRAS и не были предложены для статуса QPS (EFSA, 2007).
В целом, согласно GRAS, пробиотики признаны полностью безопасными, поскольку они достаточно хорошо переносятся человеческим организмом. Безопасным штаммом, имеющим статус GRAS, признаны бифидобактерии B. lactis BB12, они одобрены к применению Управлением по контролю за качеством пищевых продуктов и медикаментов США (Food and Drug Administration, FDA). Клетки стрептококков S. thermophilus TH4 также признаны FDA безопасным штаммом с присвоением статуса GRAS.
2 ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
2.1 ИСПОЛЬЗОВАННЫЕ МАТЕРИАЛЫ И МЕТОДЫ
2.1.1 Протокол для фрагментации дцДНК и приготовления NSG-библиотек из 5–500 нг очищенной дцДНК
Исходный материал: 5–500 нг очищенной дцДНК (например, бактериальной геномной ДНК или геномной ДНК человека).
2.1.1.1 Фрагментация дцДНК
В ПЦР-пробирку на льду внести следующие компоненты:
| Компонент | Объем, мкл |
| 10x SD-буфер | 2,5 |
| MgCl2 (50 мМ) | 1,75 |
| дцДНК (50–500 нг) | 1–5 |
| Смесь ферментов для фрагментации (SDpol+ДНКаза I) | 1 |
| ddH2O | До 25 |
Инкубировали в ПЦР-амплификаторе в следующем режиме:
| Температура, оС | Время |
| 30 | 10–60 мин |
| 70 | 20 мин |
2.1.1.2 Лигирование индексированных вилочковых адаптеров
с фрагментированной ДНК
Добавить следующие компоненты в пробирку с фрагментированной ДНК и перемешать встряхиванием.
| Компонент | Объем, мкл |
| Фрагментированная дцДНК | 25 |
| 10x T4 лигазный буфер | 5 |
| Индексированные вилочковые адаптеры* (12,5 мкM) | 2 |
| ddH2O | 16 |
| T4 ДНК-лигаза (5 Ед/мкл) | 2 |
| Примечание: * В наборе используются индексированные адаптеры Illumina. | |
Инкубировать лигазную смесь (50 мкл) в течение 20 мин при комнатной температуре. После инкубации развести лигазную смесь 0,1х ТЕ-буфером до конечного объема 100 мкл. Образцы на этом этапе можно заморозить для хранения при минус 20 °C.
Набор содержит следующие адаптеры Illumina:
| № | Индекс Illumina | Структура |
| 1 | Universal | 5’AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATC*T |
| 2 | Index 2 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACCGATGTATCTCGTATGCCGTCTTCTGCTTG |
| 3 | Index 3 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACTTAGGCATCTCGTATGCCGTCTTCTGCTTG |
| 4 | Index 4 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACTGACCAATCTCGTATGCCGTCTTCTGCTTG |
| 5 | Index 5 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACACAGTGATCTCGTATGCCGTCTTCTGCTTG |
| 6 | Index 6 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACGCCAATATCTCGTATGCCGTCTTCTGCTTG |
| 7 | Index 9 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACGATCAGATCTCGTATGCCGTCTTCTGCTTG |
| 8 | Index 10 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACTAGCTTATCTCGTATGCCGTCTTCTGCTTG |
| 9 | Index 11 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACGGCTACATCTCGTATGCCGTCTTCTGCTTG |
| 10 | Index 12 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACCTTGTAATCTCGTATGCCGTCTTCTGCTTG |
| 11 | Index 14 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACAGTTCCGTATCTCGTATGCCGTCTTCTGCTTG |
| 12 | Index 15 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACATGTCAGAATCTCGTATGCCGTCTTCTGCTTG |
| 13 | Index 16 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACCCGTCCCGATCTCGTATGCCGTCTTCTGCTTG |
2.1.1.3 Очистка ДНК, лигированной с адаптерами
Необходимо дать шарикам AMPure XP прогреться до комнатной температуры не менее 30 мин перед использованием. Для ресуспендирования встряхнуть шарики AMPure XP на вортексе.
Добавить 80 мкл (0,8X) ресуспендированных шариков к реакции лигирования адаптеров. Хорошо перемешать пипетированием по меньшей мере 10 раз. Манипуляции следует проводить осторожно, чтобы исключить всю жидкость из наконечника во время последнего смешивания. Также можно использоваться смешивание на вортексе в течение 3–5 с. Если после центрифугирования образцы центрифугируются, необходимо остановить центрифугирование до того, как шарики начнут оседать.
Инкубировать образцы на столе не менее 5 мин при комнатной температуре.
Чтобы отделить шарики от супернатанта, планшет/пробирки помещают на соответствующую магнитную подставку. При необходимости образец центрифугируют, чтобы собрать жидкость со стенок лунки пробирки или планшета перед постановкой на магнитную подставку.
Через 5 мин (или когда раствор станет прозрачным) осторожно удаляют супернатант. Это действие надо проводить осторожно, чтобы не потревожить шарики, содержащие целевую ДНК.
Добавить 200 мкл 80%-го свежеприготовленного этанола в пробирку/планшет, находящиеся в магнитной подставке. Инкубируют при комнатной температуре в течение 30 с, а затем осторожно удаляют и отбрасывают супернатант.
Повторяют эту стадию промывки один раз, чтобы получить в итоге две стадии промывки. Обязательно удалить всю видимую жидкость после второй промывки. При необходимости, кратковременно центрифугируйте пробирку/планшет, поместите обратно на магнит и удалите следы этанола с помощью автоматического дозатора на 20 мкл. Это действие надо проводить осторожно, чтобы не потревожить шарики, содержащие целевую ДНК.
Высушить шарики на воздухе до 5 мин, пока пробирка/планшет находятся на магнитной подставке с открытой крышкой.
Внимание: важно не пересушивать шарики. Это может привести к снижению выделения ДНК-мишени. Образцы следует элюировать, когда шарики остаются темно-коричневого цвета и выглядят блестящими, но когда вся видимая жидкость испарилась. Когда шарики становятся светло-коричневыми,
и их поверхность начинает покрываться трещинами, они являются уже пересушенными.
Снимите пробирку/планшет с магнитной подставки. Элюируйте ДНК-мишень с шариков, добавляя 22–23 мкл 10 мМ Трис-HCl или 0,1X TE. Хорошо перемешайте суспензию пипетированием 10 раз или встряхиванием на вортексе. Инкубируйте суспензию не менее 2 мин при комнатной температуре. При необходимости шарики можно центрифугировать в течение короткого времени, чтобы собрать жидкость со стенок пробирки или лунок планшета, прежде чем поставить их обратно на магнитную подставку.
Поместите пробирку/планшет на магнитную подставку. Через 5 мин (или когда раствор станет прозрачным), перенести 20 мкл в новую ПЦР-пробирку для амплификации.
Примечание: образцы можно хранить при минус 20 °C.
2.1.1.4 ПЦР-амплификация лигированной ДНК адаптера
Добавить следующие компоненты в новую ПЦР-пробирку:
| Компонент | Объем, мкл |
| ДНК, лигированная с адаптерами | 20 |
| Смесь праймеров для ПЦР* (20 мкM (по 10 мкМ каждого праймера) | 4 |
| 10x SD-буфер | 5 |
| MgCl2 (50 мM) | 3,5 |
| Смесь ферментов для амплификации (20 Ед/мкл) | 1 |
| ddH2O | 16,5 |
| Примечание: *ПЦР-праймеры – P5.2_PCR 5’ AATGATACGGCGACCACCGA 3’, P7.1_PCR 5’ CAAGCAGAAGACGGCATACGAGAT 3’. | |
Смешать компоненты быстрым встряхиванием на вортексе и сбросить капли центрифугированием. Поместить пробирку в термоциклер и провести ПЦР-амплификации с использованием следующих условий циклов:
| Температура, оС | Время, с | Циклы |
| 94 | 120 | 1 |
| 95 | 20 | 5–12** |
| 62 | 20 | |
| 68 | 45 | |
| 68 | 45 | 1 |
| 4 | ∞ | |
| Примечание: ** количество циклов ПЦР следует выбирать, исходя из количества введенной ДНК. | ||
Число циклов должно быть достаточно большим, чтобы обеспечить достаточное количество фрагментов библиотеки для успешного секвенирования, но также должно быть достаточно низким, чтобы избежать появления артефактов вследствие ПЦР и избыточной амплификации (появления высокомолекулярных фрагментов на Биоаналайзере). Рекомендованное количество циклов ПЦР приведено ниже. Его можно использовать в качестве отправной точки для определения числа циклов ПЦР, наилучшего для приготовления образцов библиотек ДНК.
| Количество исходной ДНК, нг | Число циклов ПЦР, требуемое для выхода библиотеки примерно 250–500 нг |
| ≥ 250 | 5 |
| 200 | 6 |
| 100 | 7 |
| 40–50 | 8–9 |
| 20 | 10 |
| 10 | 11 |
| 5 | 12 |
2.1.1.5 Очистка ПЦР-реакции
Необходимо дать шарикам AMPure XP прогреться до комнатной температуры не менее 30 мин перед использованием. Для ресуспендирования встряхнуть шарики AMPure XP на вортексе.
Добавить 45 мкл (0,9X) ресуспендированных шариков к ПЦР-реакции. Хорошо перемешать с пипетированием по меньшей мере 10 раз. Манипуляции следует проводить осторожно, чтобы исключить всю жидкость из наконечника во время последнего смешивания. Также можно использоваться смешивание на вортексе в течение 3–5 с. Если после центрифугирования образцы центрифугируются, необходимо остановить центрифугирование до того, как шарики начнут оседать.
Инкубировать образцы на столе не менее 5 мин при комнатной температуре.
Чтобы отделить шарики от супернатанта, планшет/пробирки помещают на соответствующую магнитную подставку. При необходимости образец центрифугируют, чтобы собрать жидкость со стенок лунки пробирки или планшета перед постановкой на магнитную подставку.
Через 5 мин (или когда раствор станет прозрачным) осторожно удаляют супернатант. Это действие надо проводить осторожно, чтобы не потревожить шарики, содержащие целевую ДНК.
Добавить 200 мкл 80%-го свежеприготовленного этанола в пробирку/планшет, находящиеся в магнитной подставке. Инкубируют при комнатной температуре в течение 30 с, а затем осторожно удаляют и отбрасывают супернатант.
Повторяют эту стадию промывки один раз, чтобы получить в итоге две стадии промывки. Обязательно удалить всю видимую жидкость после второй промывки. При необходимости, кратковременно центрифугируйте пробирку/планшет, поместите обратно на магнит и удалите следы этанола с помощью автоматического дозатора на 20 мкл. Это действие надо проводить осторожно, чтобы не потревожить шарики, содержащие целевую ДНК.
Высушить шарики на воздухе до 5 мин, пока пробирка/планшет находятся на магнитной подставке с открытой крышкой.
Внимание: важно не пересушивать шарики. Это может привести к снижению выделения ДНК-мишени. Образцы следует элюировать, когда шарики остаются темно-коричневого цвета и выглядят блестящими, но когда вся видимая жидкость испарилась. Когда шарики становятся светло-коричневыми,
и их поверхность начинает покрываться трещинами, они являются уже пересушенными.
Снимите пробирку/планшет с магнитной подставки. Элюируйте ДНК-мишень с шариков, добавляя 32–33 мкл 0,1X TE. Хорошо перемешайте суспензию пипетированием 10 раз или встряхиванием на вортексе. Инкубируйте суспензию не менее 2 мин при комнатной температуре. При необходимости шарики можно центрифугировать в течение короткого времени, чтобы собрать жидкость со стенок пробирки или лунок планшета, прежде чем поставить их обратно на магнитную подставку.
Поместите пробирку/планшет на магнитную подставку. Через 5 мин (или когда раствор станет прозрачным), перенести 30 мкл в новую ПЦР-пробирку для хранения при минус 20 °C.
Далее необходимо проверить распределение фрагментов библиотеки (например, с использованием Agilent Bioanalyzer High Sensitivity DNA chip).
2.1.2 Измерение концентрации ДНК с помощью PicoGreen dsDNA Assay Kit
В ходе работы не следует использовать для раствора стеклянную посуду.
Подготовка красителя:
Перед началом работы следует дать реагенту Quant-iT™ PicoGreen® нагреться до комнатной температуры, прежде чем открывать пробирку с ним. Для наилучшего результата раствор необходимо использовать в течение нескольких часов после приготовления. Приготовленный раствор с красителем следует защищать от света (помещая его в темное место или накрывая фольгой).
- Проверить корректность установки даты и времени в приборе (Settings → «Instrument settings» → «Date/Time»). При необходимости установить верные значения.
- В чистую 15-миллилитровую пробирку внести 1005 мкл 20х TE буфера.
- Добавить в пробирку 19095 мкл Н2О.
- Добавить к нему 101 мкл красителя.
- Тщательно перемешать.
- Приготовить контрольную ДНК для построения кривой (0 и 100 нг
в 5 мкл):- внести в пробирку на 0,2 мл 8 мкл 1хТЕ буфера;
- добавить 2 мкл контрольной ДНК из набора (100нг/мкл). Хорошо перемешать пипетированием;
- подготовить две оптически прозрачные пробирки на 0,5 мл;
- одну подписать как К1, другую – К2;
- в К1 внести 200 мкл раствора для измерения концентрации;
- в К2 внести 195 мкл раствора для измерения и 5 мкл разведенной контрольной ДНК (из шага b);
- Раскапать по 195 мкл в оптически прозрачные пробирки на 0,5 мл.
- Внести в каждую пробирку по 5 мкл соответствующей коровьей ДНК.
- Внести в пробирку К1 5 мкл раствора ДНК второго разведения
(38 + 2 мкл). - Внести в пробирку К2 5 мкл раствора ДНК первого разведения
(8 + 2 мкл). - Включить прибор, подключить его к компьютеру.
- Выбрать на первом экране программу PicoGreen.
- Произвести измерения.
2.1.3 Подготовка библиотеки к нанесению в картридж
После объединения образцов измерить концентрацию ДНК.
Произвести пересчет массовой концентрации в молярность, программа выдает данные в пикомолях, необходимо пересчитать в наномоли – http://molbiol.ru/scripts/01_07.html.
Далее необходимо следовать инструкциям Protocol A из «MiSeq System Denature and Dilute Libraries Guide».
2.1.3.1 Разведение щелочи (NaOH)
К 100 мкл стокового раствора (10М) добавить 900 мкл Н2О.
К отобранным 200 мкл полученного раствора (1N NaOH) добавить
800 мкл Н2О.
В результате получится 1 мл 0,2N NaOH.
2.1.3.2 Денатурация библиотеки
1. В микроцентрифужной пробирке смешать следующие объемы.
- 4 нM библиотека (5 мкл);
- 0,2N NaOH (5 мкл).
2. Перемешать кратким встряхиванием на вортексе и центрифугировать при 280×g в течение минуты.
3. Инкубировать при комнатной температуре в течение 5 мин.
4. В пробирку, содержащую денатурированную библиотеку, добавить
990 мкл холодного НТ1-буфера.
5. В результате получается 1 мл денатурированной библиотеки с концентрацией 20 пМ.
6. Нанести в картридж 600 мкл.
2.2 СОЗДАНИЕ ПРЕДСТАВИТЕЛЬНЫХ ГЕНОИНЫХ БИБЛИОТЕК ДЛЯ МАССИВНОГО ПАРАЛЛЕЛЬНОГО СЕКВЕНИРОВАНИЯ ИССЛЕДУЕМЫХ ПРОБИОТИЧЕСКИХ ШТАММОВ
2.2.1 Приготовление геномных библиотек для массивного параллельного секвенирования
Было выделено восемь образцов геномной ДНК для следующих клонов
с рабочими названиями: 1sq, 1/2sq, 2sq, 5sq, 7sq, 8sq, 10sq, MhP. Видовая принадлежность соответствующих клонов, установленная путем анализа фрагментов последовательностей генов 16S РНК, указана в таблице 7.
Таблица 7 – Видовая принадлежность образцов геномной ДНК
| 1sq | Lactobacillus salivarius |
| 1/2sq | Lactobacillus salivarius |
| 2sq | Lactobacillus curvatus |
| 5sq | Lactobacillus buchneri |
| 7sq | Lactobacillus parabuchneri |
| 8sq | Lactobacillus heilongjiangensis |
| 10sq | Lactobacillus sp. |
| MhP | Lactobacillus sp. |
2.2.2 Анализ выделенной геномной ДНК с помощью электрофореза в агарозном геле
По 5 мкл из каждого образца наносили на 1,2%-й агарозный гель, содержащий бромид этидия в концентрации 0,5 мкг/мкл, в буфере для наноса образцов и разделяли электрофорезом при напряженности электрического поля
5 В/см. Результат представлен на рисунке 3.
Рисунок 3 – Изображение образцов геномной ДНК, разделенной
электрофорезом в 1,2%-ом агарозном геле, содержащем 0,5 мкг этидий
бромида. Слева находится маркер молекулярных весов. Снизу указаны
краткие (рабочие) названия выделенных образцов ДНК, видовая
принадлежность которых указана в таблице 7
Из рисунка3 видно, что все образцы содержали геномную ДНК, однако
в разных количествах, сильно отличающихся друг от друга. Образец 8sq имел наименьшую геномной ДНК, тогда как образцы 1sq и 10sq содержали больше всего геномной ДНК. Кроме того, из рисунка видно, что образцы 1sq, 10sq
и MhP загрязнены РНК. Для очистки образцов от РНК, а также для выравнивания и концентрирования полученной геномной ДНК проводили дополнительную очистку и концентрирование геномной ДНК на магнитных шариках AMPure XP.
2.2.3 Очистка геномной ДНК на магнитных шариках AMPure XP Beads
Для очистки шарикам AMPure XP давали прогреться до комнатной температуры не менее 30 мин перед использованием. Шарики AMPure XP Beads разбивали на вортексе для ресуспендирования. Добавляли 30 мкл (0,6X) ресуспендированных шариков для процедуры очистки ДНК. Хорошо перемешивали шарики с ДНК пипетированием с использованием автоматического дозатора по крайней мере 10 раз. Инкубировали смешанные с шариками образцы ДНК на столе 5 мин при комнатной температуре. После этого капли со стенок собирали кратким (15 с) центрифугированием при 8000g. Далее пробирки, содержащие образцы ДНК с шариками, помещали на магнитную подставку, чтобы отделить шарики от супернатанта. Инкубировали смесь ДНК с шариками на магнитной подставке в течение 5 мин и аккуратно удаляли надосадочную жидкость. Шарики промывали 2 раза, добавляя по 200 мкл 80%-го свежеприготовленного этанола в пробирку на магнитной подставке. Инкубировали при комнатной температуре в течение 30 с, а затем осторожно удаляли надосадочную жидкость. После второй промывки удаляли остаточную жидкость с помощью автоматического дозатора на 20 мкл. Шарики высушивали на воздухе в течение
5 мин, при этом пробирка оставалась на магнитной подставке. ДНК с шариков элюировали 30 мкл 0,1х ТЕ-буфера. 5 мкл элюата наносили на 1,2%-й агарозный гель, содержащий 0,5 мкг/мл бромистого этидия, и разделяли электрофорезом при напряженности электрического поля 5 В/см.
Результаты разделения очищенных образцов ДНК приведены на рисунке 4.
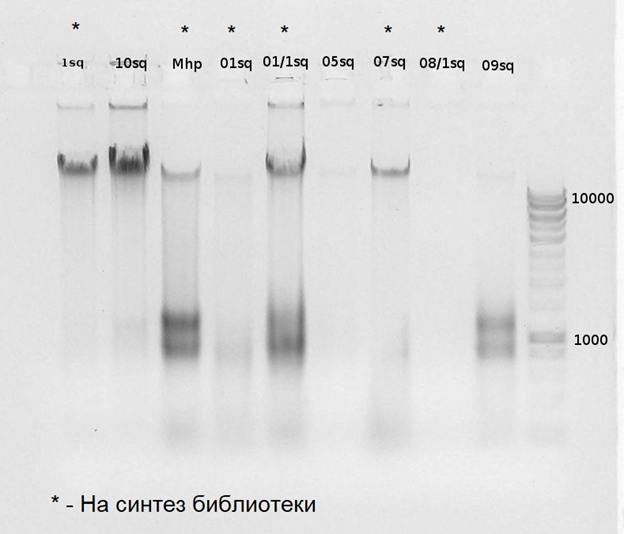
Рисунок 4 – Изображение образцов геномной ДНК, разделенной
электрофорезом при 5 В/см в 1,2%-ом агарозном геле, содержащем 0,5 мкг
этидий бромида. Маркер молекулярных весов находится справа. Сверху
указаны названия прошедших очистку на магнитных шариках
AMPure XP Beads образцов геномной ДНК
Из рисунка 4 видно, что после проведения процедуры очистки/нормализации ДНК на шариках AMPure XP Beads содержание геномной ДНК в образцах значительно выровнялось, а кроме того на существенном уровне была удалена РНК из образцов.
Однако для приготовления библиотек кроме качественной характеристики выделенной ДНК также было необходимо определить точную концентрацию выделенной геномной ДНК.
2.2.4 Определение концентрации геномной ДНК с помощью красителя PicoGreen
Концентрацию ДНК определяли с помощью набора Quant-iT PicoGreen dsDNA Assay Kit (Thermo Fisher Scientific), следуя инструкциям производителя. Этот набор, основанный на методе с использованием красителя PicoGreen, выгодно отличается от всех других способов измерения, тем что данный краситель высокоизбирательно связывается с двухцепочечной ДНК, при этом не окрашивая одноцепочечную ДНК или РНК, что позволяет использовать его для высокоточных измерений концентрации ДНК в присутствии РНК. Кроме того, этот краситель позволяет измерять концентрации ДНК в широком диапазоне, включая пикомолярный. Таким образом, для определения концентрации геномной ДНК в образцах было принято решение использовать данный метод с красителем PicoGreen.
Для этого приготавливали свежий раствор красителя PicoGreen в разведении 1 : 200 в ТЕ-буфере. После этого приготавливали необходимое количество оптически прозрачных пробирок, в которые добавляли по 200 мкл раствора PicoGreen в ТЕ-буфере. Для построения калибровочной кривой использовали ДНК фага лямбда в ТЕ. Калибровочную кривую строили с использованием 2–
5 концентраций ДНК фага лямбда. Концентрацию образцов ДНК определяли, добавляя в 200 мкл раствора PicoGreen в ТЕ-буфере по 5 мкл раствора измеряемого образца (фактор разведения 40). Конечную измеряемую концентрацию
и концентрацию ДНК с учетом фактора разведения определяли с использованием программы, записанной в памяти прибора.
В таблице 8 приведены концентрации геномной ДНК, измеренной с помощью набора Quant-iT PicoGreen dsDNA Assay Kit.
Таблица 8 – Концентрации гемномной ДНК
| Номер | Рабочее название | Концентрация выделенной ДНК | Единицы |
| 1 | 1sq | 48 | нг/мкл |
| 2 | 1/2sq | 4,72 | нг/мкл |
| 3 | 2sq | 2,66 | нг/мкл |
| 5 | MhP | 66,4 | нг/мкл |
| 6 | 01sq | 27,96 | нг/мкл |
| 7 | 01/1sq | 70,8 | нг/мкл |
| 9 | 07sq | 34,04 | нг/мкл |
| 10 | 08/1sq | 0,704 | нг/мкл |
Из таблицы 8 видно, что концентрации выделенной геномной ДНК отличаются друг от друга, но все (за исключением образца № 10, 08sq) подходят для концентрационных диапазонов, указанных в наборе для приготовления геномных библиотек. Тем не менее было решено также использовать образец 08sq для создания библиотеки, поскольку несмотря на низкую концентрацию он все-таки содержал геномную ДНК, и было решено проверить возможность использования набора для создания библиотек с использованием геномной ДНК
в концентрациях, значительно ниже указанной в описании к набору.
2.2.5 Синтез библиотек для массивного параллельного секвенирования (NSG)
Исходя из данных качества выделенной и очищенной геномной ДНК, полученных электрофорезом в агарозном геле, и концентрации, полученной флуоресцентным методом с использованием красителя PicoGreen, было решено для синтеза использовать количество геномной ДНК и число циклов амплификации, которые приведены в таблице 9.
2.2.6 Фрагментация ДНК
На льду размораживали компоненты набора, используемые для фрагментации ДНК. На льду приготавливали для каждого образца ДНК смесь из компонентов набора. Количества указаны в таблице 10.
Таблица 9 – Количество геномной ДНК, использованной для синтеза
NSG-библиотек, и число циклов, используемых в процессе
амплификации при приготовлении NSG-библиотек
| Номер | Рабочее название |
мкл на синтез |
нг на синтез |
мкл воды до 5 мкл |
Наложенный циклеж |
| 1 | 1sq | 4 | 200 | 1 | 6 |
| 2 | 1/2sq | 5 | 20 | 0 | 14 |
| 3 | 2sq | 5 | 15 | 0 | 14 |
| 5 | MhP | 3 | 200 | 2 | 6 |
| 6 | 01sq | 5 | 150 | 0 | 6 |
| 7 | 01/1sq | 3 | 200 | 2 | 6 |
| 9 | 07sq | 5 | 150 | 0 | 6 |
| 10 | 08/1sq | 5 | 5 | 0 | 14 |
Таблица 10 – Схема приготовления смеси для фрагментации геномной ДНК
| Компоненты | Объем, мкл |
| 10x SD-буфер | 2,5 |
| MgCl2 (50 мM) | 1,75 |
| Смесь ферментов для фрагментации (SDpol и ДНКаза I) | 1 |
| ddH2O | До 20 |
К полученной смеси добавляли указанные в таблице 9 количества геномной ДНК и воды, сбрасывали на центрифуге, смешивали и снова сбрасывали, и проводили процедуру фрагментации ДНК согласно протоколу, указанному в таблице 11. Процедуру фрагментации геномной ДНК проводили в ПЦР-амплификаторе. Стадия прогрева при 70 °С была необходима для инактивации ДНКазы I.
Таблица 11 – Протокол фрагментации геномной ДНК
| Температура, оС | Время |
| 30 | 20 мин |
| 70 | 20 мин |
После фрагментации образцы помещали на лед до проведения следующей процедуры.
2.2.7 Лигирование индексированных вилочковых адаптеров в фрагментированной ДНК
К полученным фрагментам геномной ДНК с помощью процесса лигирования присоединяли предварительно гибридизованные пары индексированных вилочковых адаптеров и универсального адаптера.
В таблице 12 приведены последовательности универсального адаптера
и адаптеров, используемых для синтеза NSG-библиотек для каждого образца.
Таблица 12 – Нуклеотидные последовательности адаптеров, используемых
в синтезе NSG-библиотек
| Номер | Рабочий номер | Индекс Illumina | Структура |
| Универсальный | 5’AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATC*T | ||
| 1 | 1sq | Index 2 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACCGATGTATCTCGTATGCCGTCTTCTGCTTG |
| 2 | 1/2sq | Index 3 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACTTAGGCATCTCGTATGCCGTCTTCTGCTTG |
| 3 | 2sq | Index 4 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACTGACCAATCTCGTATGCCGTCTTCTGCTTG |
| 4 | MhP | Index 5 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACACAGTGATCTCGTATGCCGTCTTCTGCTTG |
| 5 | 01sq | Index 6 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACGCCAATATCTCGTATGCCGTCTTCTGCTTG |
| 6 | 01/1sq | Index 9 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACGATCAGATCTCGTATGCCGTCTTCTGCTTG |
| 7 | 07sq | Index 10 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACTAGCTTATCTCGTATGCCGTCTTCTGCTTG |
| 8 | 08/1sq | Index 11 | 5’PGATCGGAAGAGCACACGTCTGAACTCCAGTCACGGCTACATCTCGTATGCCGTCTTCTGCTTG |
В пробирку с фрагментированной ДНК на льду добавляли компоненты набора согласно количествам, приведенным в таблице 13.
Таблица 13 – Схема приготовления смеси для проведения реакции
лигирования пар индексированных адаптеров
| Компонент | Объем, мкл |
| 10x T4 лигазный буфер | 5 |
| Гибридизованные пары адапторов (12,5 мкM) | 2 |
| ddH2O | 16 |
| ДНК-лигаза фага Т4 (5 Ед/мкл) | 2 |
Полученную смесь (объем 50 мкл) собирали центрифугированием, перемешивали встряхиванием, собирали центрифугированием и инкубировали
20 мин при комнатной температуре – 20–22 °С.
После инкубации разводили лигированную смесь 1 : 1 0,1х ТЕ-буфером до конечного объема 100 мкл.
2.2.8 Очистка фрагментированной геномной ДНК с прикрепленными адаптерами на магнитных шариках AMPure XP Beads
Для очистки шарикам AMPure XP давали прогреться до комнатной температуры не менее 30 мин перед использованием. Шарики AMPure XP Beads разбивали на вортексе для ресуспендирования.
К 100 мкл смеси фрагментов геномной ДНК с лигированными парами индексированных адаптеров добавляли по 80 мкл (0,8X) ресуспендированных шариков для процедуры очистки ДНК. Хорошо перемешивали шарики с ДНК встряхиванием на вортексе в течение 5–10 с. Инкубировали смешанные с шариками образцы ДНК на столе 5 мин при комнатной температуре. После этого капли со стенок собирали кратким (15 с) центрифугированием при 8000g. Далее пробирки, содержащие образцы ДНК с шариками, помещали на магнитную подставку, чтобы отделить шарики от супернатанта. Инкубировали смесь ДНК с шариками на магнитной подставке в течение 5 мин и аккуратно удаляли надосадочную жидкость. Шарики промывали 2 раза, добавляя по 200 мкл 80%-го свежеприготовленного этанола в пробирку на магнитной подставке. Инкубировали при комнатной температуре в течение 30 с, а затем осторожно удаляли надосадочную жидкость. После второй промывки удаляли остаточную жидкость с помощью автоматического дозатора на 20 мкл. Шарики высушивали на воздухе в течение 5 мин, при этом пробирка оставалась на магнитной подставке. ДНК с шариков элюировали в 20 мкл 0,1х ТЕ-буфера и сразу использовали
в последующей реакции ПЦР-амплификации.
2.2.9 Амплификация смеси фрагментов геномной ДНК с лигированными парами индексированных адаптеров
В новых ПЦР-пробирках приготавливали следующую амплификационную смесь для каждого образца геномной ДНК. Состав смеси приведен в таблице 14.
В таблице 15 приведена структура используемых для ПЦР праймеров.
Таблица 14 – Схема приготовления смеси для ПЦР-амплификации смеси фрагментов геномной ДНК с лигированными парами
индексированных адаптеров
| Компонент | Объем, мкл |
| Смесь праймеров для ПЦР (20 мкM (10 мкМ каждого)) | 4 |
| 10x SD-буфер | 5 |
| MgCl2 (50 mM) | 3,5 |
| Амплификационная смесь ферментов (20 Ед/мкл) | 1 |
| ddH2O | 16,5 |
Таблица 15 – Нуклеотидная последовательность праймеров, используемых
для амплификации
| Название праймера | Последовательность |
| P5.2_PCR | 5’ AATGATACGGCGACCACCGA 3’ |
| P7.1_PCR | 5’ CAAGCAGAAGACGGCATACGAGAT 3’ |
К смеси компонентов для проведения ПЦР добавляли элюированные образцы ДНК. Смешивали компоненты для ПЦР встряхиванием и собирали центрифугированием. После чего пробирки помещали в амплификатор и проводил реакцию ПЦР при параметрах, указанных в таблице 16. При этом для каждого образца использовали число циклов ПЦР, приведенное в таблице 9.
Таблица 16 – Параметры проведения реакции амплификации фрагментов
геномной ДНК, лигированных с парами индексированных
адаптеров
| Температура, оС | Время, с | Число циклов |
| 94 | 120 | 1 |
| 95 | 20 | 6 или14 (см. таблицу 3) |
| 62 | 20 | |
| 68 | 45 | |
| 68 | 45 | 1 |
| 4 | ∞ |
Число циклов выбирали, исходя из количества вводимой ДНК. Соответственно при количестве исходной ДНК 100–200 нг использовали 6 циклов,
а при количестве исходной ДНК 5 нг – использовали 14 циклов амплификации.
2.2.10 Очистка амплифицированных библиотек после ПЦР-реакции на магнитных шариках AMPure XP Beads
Для очистки шарикам AMPure XP давали прогреться до комнатной температуры не менее 30 мин перед использованием. Шарики AMPure XP Beads разбивали на вортексе для ресуспендирования.
К 50 мкл аплифицированных с помощью ПЦР библиотек добавляли по
45 мкл (0,9X) ресуспендированных шариков для процедуры очистки ДНК. Хорошо перемешивали шарики с ДНК встряхиванием на вортексе в течение 5–
10 с. Инкубировали смешанные с шариками образцы ДНК на столе 5 мин при комнатной температуре. После этого капли со стенок собирали кратким (15 с) центрифугированием при 8000g. Далее пробирки, содержащие образцы ДНК
с шариками, помещали на магнитную подставку, чтобы отделить шарики от супернатанта. Инкубировали смесь ДНК с шариками на магнитной подставке
в течение 5 мин и аккуратно удаляли надосадочную жидкость. Шарики промывали 2 раза, добавляя по 200 мкл 80%-го свежеприготовленного этанола в пробирку на магнитной подставке. Инкубировали при комнатной температуре
в течение 30 с, а затем осторожно удаляли надосадочную жидкость. После второй промывки удаляли остаточную жидкость с помощью автоматического дозатора на 20 мкл. Шарики высушивали на воздухе в течение 5 мин, при этом пробирка оставалась на магнитной подставке. ДНК с шариков элюировали
в 30 мкл 0,1х ТЕ-буфера.
5 мкл элюированной ДНК геномных библиотек для NSG-секвенирования использовали для оценки качества и размера полученных библиотек. Концентрацию ДНК библиотек измеряли с помощью набора Quant-iT PicoGreen dsDNA Assay Kit.
Оценка качества и размера полученных NSG-библиотек электрофорезом
в 1,2%-ом агарозном геле.
Качество и размер полученных NSG-библиотек оценивали с помощью электрофореза в 1,2%-ом агарозном геле, содержащем 0,5 мкг/мл бромистого этидия, при напряженности электрического поля 5 В/см. Для этого использовали 5 мкл элюированной ДНК. Результаты разделения показаны на рисунке 5
(А и Б). Панель А рисунка 5 соответствует короткому прогону разделяемой ДНК, а панель Б – длинному прогону разделяемой ДНК.
А)
Б)
Рисунок 5 – Изображение образцов геномной ДНК, разделенной
электрофорезом при 5 В/см в 1,2%-ом агарозном геле, содержащем 0,5 мкг
этидий бромида. Маркер молекулярных весов находится справа. Сверху
указаны названия образцов, используемых для создания библиотек
Из результатов качественной оценки видно, что все образцы соответствуют требуемому разбросу размеров фрагменов и концентраций. Однако поскольку все-таки наблюдается некоторая вариабельность в концентрациях синтезированных библиотек, концентрацию ДНК в них далее определяли количественно и приготавливали смесь библиотек исходя из количественной оценки концентраций библиотек.
2.2.11 Определение концентрации NSG-библиотек с помощью красителя PicoGreen
Концентрацию ДНК в синтезированных NSG-библиотеках определяли
с помощью набора Quant-iT PicoGreen dsDNA Assay Kit (Thermo Fisher Scientific), следуя инструкциям производителя.
Для этого приготавливали свежий раствор красителя PicoGreen в разведении 1 : 200 в ТЕ-буфере. После этого приготавливали необходимое количество оптически прозрачных пробирок, в которые добавляли по 200 мкл раствора PicoGreen в ТЕ-буфере. Для построения калибровочной кривой использовали ДНК фага лямбда в ТЕ. Калибровочную кривую строили с использованием 2–
5 концентраций ДНК фага лямбда. Концентрацию образцов ДНК определяли, добавляя в 200 мкл раствора PicoGreen в ТЕ-буфере по 5 мкл раствора измеряемого образца (фактор разведения 40). Конечную измеряемую концентрацию
и концентрацию ДНК с учетом фактора разведения определяли с использованием программы, записанной в памяти прибора.
В таблице 17 приведены концентрации ДНК в синтезированных NSG-библиотеках, измеренные с помощью набора Quant-iT PicoGreen dsDNA Assay Kit.
Таблица 17 – Концентрации ДНК в синтезированных NSG-библиотеках
| Номер | Рабочее название | Концентрация ДНК | Единицы |
| 1 | 1sq | 4,2 | нг/мкл |
| 2 | 1/2sq | 4 | нг/мкл |
| 3 | 2sq | 2,024 | нг/мкл |
| 5 | MhP | 2,516 | нг/мкл |
| 6 | 01sq | 1,08 | нг/мкл |
| 7 | 01/1sq | 6,56 | нг/мкл |
| 9 | 07sq | 2,152 | нг/мкл |
| 10 | 08/1sq | 0,536 | нг/мкл |
Таким образом, на данном этапе исполнения проекта получено достаточное количество тотальной геномной ДНК исследуемых штаммов и созданы
в необходимой концентрации библиотеки для секвенирования бактериальных геномов отобранных образцов с использованием набора MiSeq Reagent Micro Kit v2 (300-cycles) MS-103-1002 для получения наибольшего покрытия, на платформе Illumina MiSeq.
2.3 РЕЗУЛЬТАТЫ АНАЛИЗА ПОЛНОГЕНОМНЫХ СИКВЕНСОВ ПРОБИОТИЧЕСКИХ ШТАММОВ
За отчетный период (2019 г.) получены чистые культуры штаммов
L. salivarius PC20b и L. curvatus PC723_5. Выделены тотальные геномные ДНК этих культур, и получены очищенные нормализованные образцы. Впервые двумя независимыми методами созданы представленные геномные библиотеки для массивного параллельного секвенирования (NGS). Осуществлено секвенирование подготовленных библиотек на платформе Illumina MiSeq. В ходе необходимого числа запусков секвенирования получены массивы данных, содержащие информацию о фрагментах генома штаммов L. salivarius PC20b и L. curvatus PC723_5, которые позволили восстановить полногеномные последовательности и уточнить таксономическую идентификацию штаммов. В результате сравнительного биоинформационного анализа собраны четыре генома, депонированные в международную базу данных NCBI.
Проведен предварительный структурный анализ геномов депонированных штаммов. Полученные данные послужат основой для выяснения молекулярных механизмов реализации пробиотических и антагонистических свойств исследуемых штаммов L. salivarius PC20b и L. curvatus PC723_5. Кроме того, проведенные в отчетном году исследования и приобретенные коллективом компетенции обеспечивают успешность реализации планов последующих лет и выполнение поставленных в проекте задач.
2.3.1 Депонирование полногеномных сиквенсов в международную базу данных NCBI
Все цели, поставленные на 2019 г., достигнуты в полной мере. Более того, в ходе полногеномного секвенирования штаммов L. salivarius PC20b и L. curvatus PC723_5 проведено уточнение их таксономической принадлежности, собраны последовательности геномов этих штаммов. Полногеномные последовательности подготовлены к депонированию и размещены в международной базе NCBI (англ. National Center for Biotechnological Information – Национальный центр биотехнологической информации США) с присвоением соответствующих индивидуальных номеров.
1. На основании выделенных ранее чистых бактериальных культур штаммов L. salivarius PC20b и L. curvatus PC723_5 получено достаточное количество биомассы для выделения тотальной геномной ДНК.
2. При помощи коммерчески доступного набора Genomic DNA Purification Kit (#K0512, Thermo Scientific, USA) получены образцы тотальной геномной ДНК штаммов L. salivarius PC20b и L. curvatus PC723_5. ДНК очищена и нормализована при помощи магнитных шариков AMPure XP Beads (Thermo Scientific, USA).
3. Определена концентрация ДНК с помощью набора Quant-iT PicoGreen dsDNA Assay Kit (Thermo Fisher Scientific), с использованием красителя PicoGreen.
4. Синтезировано шесть библиотек (по три каждого из изучаемых штаммов) для массивного параллельного секвенирования (NSG).
5. Осуществлено секвенирование бактериальных геномов отобранных образцов с использованием набора MiSeq Reagent Micro Kit v2 (300-cycles) MS-103-1002 для получения наибольшего покрытия, на платформе Illumina MiSeq.
6. С использованием автоматической платформы Illumina MiSeq осуществлена сборка геномов и проведено сравнение с аннотированными референсными геномы, доступными в NCBI.
7. Уточнено таксономическое положение штаммов L. salivarius PC20b
и L. curvatus PC723_5.
8. Проведено размещение в международной базе данных NCBI четырех последовательностей полных геномов штаммов L. salivarius PC20b и L. curvatus PC723_5. После проверки и аннотации рецензентами NCBI депонированным последовательностям присвоены индивидуальные номера:
Lactobacillus salivarius PC20b (VRA_1sq_f) (Assembly: GCA_009683065.1),
Lactobacillus salivarius PC20b (VRA_01-1sq_f) (Assembly: GCA_009683115.1),
Lactobacillus curvatus PC723_5 (VRA_2sq_f) (Assembly: GCA_009683015.1),
Lactobacillus curvatus PC723_5 (VRA_2sq_n) (Assembly: GCA_009683145.1).
9. Проведен предварительный структурный анализ геномов депонированных штаммов, который позволил определить для каждого из трех геномов GC%-состав, идентифицировать гены, псевдогены, белки и плазмиды.
Данные о полных геномах исследуемых штаммов L. salivarius PC20b
и L. curvatus PC723_5 получены впервые. Кроме того, в обоих случаях уточнена таксономическая идентификация и получены приоритетные данные о первичных последовательностях генов и белков, способных определять пробиотические и антагонистические свойства исследуемых штаммов.
Впервые получены приведенные в таблице 18 данные.
Таблица 18 – Геномы L. salivarius и L. curvatus, депонированные в NCBI (PRJNA588983)
| Штамм | Размер генома (Mb) |
GC% | Кол-во генов | Кол-во белков | Кол-во плазмид |
| L. salivarius PC20b VRA_1sq_f | 2.28982 | 32,60 | 3186 | 2886 | 30 |
| L. salivarius PC20b VRA_01-1sq_f | 2.22963 | 32,40 | 2953 | 2722 | 5 |
| L. curvatus PC723_5 VRA_2sq_f | 2.02224 | 41,20 | 2208 | 2114 | 5 |
| L. curvatus PC723_5 VRA_2sq_n | 1.97745 | 41,10 | 2034 | 1948 | 5 |
2.3.2 Анализ полученных данных о полногеномных последовательностей пробиотических штаммов
Выделение чистых культур проводили с использованием коммерчески доступных селективных сред для видов рода Lactobacillus (Лактобакагар и MRS). Предварительное таксономическое определение проводили путем определения первичной нуклеотидной последовательности фрагментов вариантов гена 16S РНК методом прямого ферментативного секвенирования по Сенгеру.
Выделение тотальной геномной ДНК проводили при помощи коммерчески доступного набора Genomic DNA Purification Kit (#K0512, Thermo Scientific, USA) согласно рекомендациям производителя. ДНК очищена и нормализована при помощи магнитных шариков AMPure XP Beads (Thermo Scientific, USA).
Определение концентрации ДНК проводили с помощью набора Quant-iT PicoGreen dsDNA Assay Kit (Thermo Fisher Scientific), с использованием красителя PicoGreen. Приготовление геномных библиотек для массивного параллельного секвенирования (NSG) проводили с помощью набора Nextera DNA Flex.
Секвенирование бактериальных геномов отобранных образцов осуществляли с использованием набора MiSeq Reagent Micro Kit v2 (300-cycles) MS-103-1002 для получения наибольшего покрытия, на платформе Illumina MiSeq. Сборку геномов осуществляли с использованием автоматической платформы Illumina MiSeq. Было проведено сравнение с аннотированными референсными геномы, доступными в NCBI.
В результате нашей работы было проведено секвенирование геномов штаммов бактерий, принадлежащих к видам Lactobacillus salivarius, Lactobacillus curvatus, Lactobacillus parabuchneri и Lactobacillus heilongjiangensis. При этом секвенирование некоторых штаммов было выполнено несколько раз.
По итогам, количество информации, полученное в первое секвенирование было разным для разных штаммов (таблица 19) и сильно разнилось. Как предполагается, это было вызвано количеством ДНК, использованным для секвенирования.
Таблица 19 – Количество информации, полученное при секвенировании
исследуемых штаммов бактерий
| Название вида | Количество информации, Кб |
| Lactobacillus salivarius | 1476745 |
| Lactobacillus curvatus | 352506 |
| Lactobacillus parabuchneri | 1160156 |
| Lactobacillus heilongjiangensis | 516300 |
| Итого: | 3505707 |
Как уже было сказано, разное количество ДНК привело к разному количество полученной информации. Тем не менее, по всем полученным геномам нами были проведены сборки геномов с использованием референсных геномов, имеющихся в базе данных NCBI (NC_018610.1 Lactobacillus buchneri CD034, NZ_CP035110.1 Lactobacillus curvatus strain SRCM103465, NZ_CP012559.1 Lactobacillus heilongjiangensis strain DSM 28069, NZ_CP018796.1 Lactobacillus parabuchneri strain FAM21731 и NC_007929.1 Lactobacillus salivarius UCC118), при помощи программы UGENE Unipro с использованием различных алгоритмов (BWA и Bowtie2). При этом по результатам сравнения сборок геномов было принято использовать алгоритм BWA.
Таким образом, нами были получены сборки некоторых геномов, которые могли быть выравнены, используя референсные последовательности. В виду того, что количество данных по некоторым штаммам не давало возможность полностью выравнить геномы и, таким образом, собрать полные их последовательности, нами было произведено повторное секвенирование (таблица 20).
Таблица 20 – Количество информации, полученное при секвенировании
исследуемых штаммов бактерий
| Название вида | Количество информации, Кб |
| Lactobacillus salivarius 01-1sq | 160172 |
| Lactobacillus salivarius 1-2sq | 3494894 |
| Lactobacillus salivarius 1sq | 130251 |
| Lactobacillus curvatus 2sq | 1753077 |
| Lactobacillus parabuchneri 07sq | 80128 |
| Lactobacillus heilongjiangensi 08-1sq | 1848677 |
| Итого: | 7467199 |
Как мы можем видеть, количество информации собранной о строении геномов бактериальных штаммов разнилось и во второй раз. При этом, как и ранее, наибольшее количество информации было получено о штамме Lactobacillus salivarius, в то время как по Lactobacillus parabuchneri – нет. Тем не менее, получилось собрать информацию о геномах Lactobacillus curvatus и Lactobacillus heilongjiangensi. Не смотря на это, количество информации, собранное о других видах бактерий было достаточно небольшим. Тем не менее, количество информации, собранное во второй раз было почти в два раз больше, чем в первый.
В виду того, что по литературным данным в отношении создания пробиотических препаратов наиболее приоритетными являются штаммы Lactobacillus salivarius 1-2sq, Lactobacillus curvatus 2sq и Lactobacillus heilongjiangensi 08-1sq, нами было проведено секвенирование этих генотипов отдельно в третий раз (таблица 21).
В итоге, после сбора данных о строении геномов, нами было проведено выравнивание исследуемых образцов на референсные последовательности. Результаты представлены на рисунках 6–10. В случае, когда геномы плохо выравнивались на референсные, для подтверждения их видовой принадлежности, нами был выполнен поиск и сравнение 16S rRNA последовательностей с референсными для этого вида бактерий, депонированными в базу данных NCBI.
Таблица 21 – Количество информации, полученное при секвенировании
исследуемых штаммов бактерий
| Название вида | Количество информации, Кб |
| Lactobacillus salivarius 1-2sq | 346172 |
| Lactobacillus curvatus 2sq | 421439 |
| Lactobacillus heilongjiangensi 08-1sq | 198894 |
| Итого: | 966505 |
1sq_S3_L001 Lactobacillus salivarius
Рисунок 6 – Результаты сравнения рефернсного генома
Lactobacillus salivarius с выравненным
Как мы можем видеть, при выравнивании в NCBI BLAST было выявлено, что длина собранного генома без «пробелов» составила 1588285 п.н., против 1827111 п.н. референсного генома.
При сравнении референсного и выравненого геномов Lactobacillus curvatus было выявлено, что длина сборки составила1203496 п.н. против 2042320 п.н. референсного. Поэтому, для уточнения систематического положения данного штамма нами был проведен поиск последовательности 16S rRNA и сравнение ее с референсной (рисунок 7).
Как мы можем видеть, сравнение последовательности 16S rRNA изучаемого генома с референсной NR_042437.1 показало 99 % схожести. В свою очередь, это дает предположить верное систематическое определение данного штамма бактерий.
2sq_S4_L001 Lactobacillus curvatus
Рисунок 7 – Результаты сравнения рефернсного генома
Lactobacillus curvatus с выравненным
Рисунок 8 – Результаты сравнения 16S rRNA последовательностей Lactobacillus salivarius
Как мы можем видеть, сборка генома штамма Lactobacillus parabuchneri прошла достаточно успешно и практически полный геном был собран
(2600578 п.н. референсного против 2539591 п.н. собранного).
Удивительным было то, что не смотря на большое количество полученных данных, нами не был собран геном бактерии Lactobacillus heilongjiangensis. Поэтому, мы также как и в прошлый раз провели сравнение 16S rRNA изучаемого штамма с уже известными и имеющимися в базе данных NCBI.
07sq_S5_L001 Lactobacillus parabuchneri
Рисунок 9 – Результаты сравнения рефернсного генома
Lactobacillus parabuchneri с выравненным
При этом интересным было то, что при сравнение генома с «пробелами»
с NR_109370.1, последовательность 16S rRNA была найдена. Поэтому, для проверки нами был проведен поиск 16S rRNA в консенсус-последовательности генома без «пробелов» (рисунок 10).
Рисунок 10 – Результаты сравнения 16S rRNA последовательностей
Lactobacillus parabuchneri
В итоге, последовательность была обнаружена с 99 % идентичности при покрытии 96 % длины сиквенса. Это, в свою очередь, дает сделать предположение о том, что вид определен правильно.
Таким образом, из желудочно-кишечного тракта дикого кавказского фазана (Phasianus colchicus), добытого на территории Краснодарского края выделены чистые культуры кисломолочных бактерий. По результатам анализа последовательностей вариантов гена 16S РНК эти культуры были отнесены к четырем видам рода Lactobacillus – потенциальным пробиотикам. По результатам полногеномного секвенирования пробиотических штаммов Lactobacillus Salivarius PC20b, Lactobacillus curvatus PC723_5, Lactobacillus parabuchneri и Lactobacillus heilongjiangensis получен большой массив данных, первичный анализ которых позволил уточнить таксономическое положение штаммов. Так за несколько циклов запуска NGS в общей сложности получено 11939411 Кб данных. Данные о полных геномах исследуемых штаммов получены впервые. Кроме того, во всех случаях уточнена таксономическая идентификация и получены приоритетные данные о первичных последовательностях генов и белков, способных определять пробиотические и антагонистические свойства исследуемых штаммов.
2.4 ИДЕНТИФИЦИРОВАННЫЕ БАКТЕРИОЦИНЫ И РЕЗУЛЬТАТЫ БИОИНФОРМАЦИОННОГО АНАЛИЗА ИХ СТРУКТУР
2.4.1 Идентификация последовательностей бактериоцинов, найденных в ходе автоматичего анализа полногеномных сиквенсов штаммов L.salivarius VRA_1sq_f и VRA_01-1sq_f
В штамме Lactobacillus salivarius PC20b впервые идентифицированы и депонированы в базу данных NCBI последовательности девяти бактериоцинов (таблица 22).
Таблица 22 – Список бактериоцинов, обнаруженных в штаммах
L. salivarius VRA_1sq_f и VRA_01-1sq_f
| Номер | Название белка | Номер в NCBI | Длина, а.о. |
| 1 | bacteriocin | MSE07706.1 | 44 |
| 2 | bacteriocin immunity protein | MSE08628.1 | 103 |
| 3 | bacteriocin | MSE08789.1 | 72 |
| 4 | bacteriocin immunity protein, partial | MSE08809.1 | 111 |
| 5 | bacteriocin | MSE08900.1 | 57 |
| 6 | bacteriocin | MSE08901.1 | 85 |
| 7 | bacteriocin | MSE08902.1 | 79 |
| 8 | bacteriocin | MSE08971.1 | 39 |
| 9 | bacteriocin immunity protein, partial | MSE09644.1 | 55 |
Ниже приведены первичные аминокислотные последовательности бактериацинов, найденных в ходе выполнения проекта и идентифицированных в ходе автоматического анализа полногеномных сиквенсов штаммов L. salivarius VRA_1sq_f и VRA_01-1sq_f.
1. MSE07706.1
MDNCNNFTSL NNTELQGIIG GKHGLGYHIV DAVVSFGEGF LNAI
2. MSE08628.1
MEKVQELQRL VVELYDSFEN DNRKGVEEIR QVLANVNEKY KTSSHPLAWT GRLVLYLQVN AVKKDLYLTP EQKDIIKELA RIGKRTNLNY VYLSPVDDAK QFV
3. MSE08789.1
MDKEFTVLTE CELAKVDGGK GSKGSSYVAG FASAAIADTG LGGAICGVPC AMIGAHYAPI GWTIVTGATG GF
4. MSE08809.1
MKKVNVEETK EVIHELYNNL MDRPQKTSGL LDITDVLLQV YKKIDTTKYP EYLLNKLVNY IYSVGFDQKI RFMGRDGELL RKLADESNKA GLNSRYRADY SAKSQFYSYS E
5. MSE08900.1
MNNNFVQIDK KELAHIIGGR NSYDYIDSGK FGYDIGCTIA NTKFFKRLRH SNQNICS
6. MSE08901.1
MLKKLWETWL DGGFIRGKKR YVIAPVIWAL LISLGIWLFG NEEMSYLDYI QTPKMIIVTI FCLVGGSTLL YLLDTAMKVN KHMKG
7. MSE08902.1
MKEFTVLTEC ELAKVDGGYT PKNCAMAVGG GMLSGAIRGG VSGTVFGVGT GNLTGAFAGA HIGLVAGGLA CIGGYLGSH
8. MSE08971.1
MKFEVLTEKK LQVVVGGTTK NFGKSQYKKC LFNFFGTCK
9. MSE09644.1
MERTKLINQA QTDIKELLGI LNNYEKKQSE LLDIIDVLAQ VYRKLPETKN PEALL
2.4.2 Биоинформационный анализ бактериоцинов. Идентификация структурных элементов
На рисунках 11‒19 приведены примеры результатов расчетов информационных структур бактериоцинов, обнаруженных нами в штаммах L. salivarius VRA_1sq_f и VRA_01-1sq_f и депонированных в международную базу данных NCBI. В подписях к рисункам указаны номера депонирования в базе в соответствии с таблицей 22.
 |
 |
 |
| Рисунок 11 – MSE07706.1 | Рисунок 12 – MSE08628.1 | Рисунок 13 – MSE08789.1 |
 |
 |
 |
| Рисунок 14 – MSE08809.1 | Рисунок 15 – MSE08900.1 | Рисунок 16 – MSE08901.1 |
 |
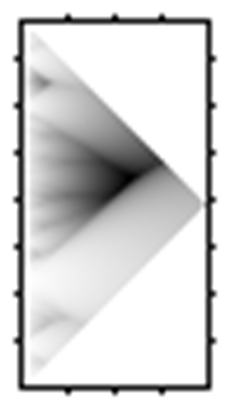 |
 |
| Рисунок 17 – MSE08902.1 | Рисунок 18 – MSE08971.1 | Рисунок 19 – MSE09644.1 |
На рисунках 11‒19 оси представляют номера остатков последовательности (по вертикали) и коэффициент сглаживания а (по горизонтали). Ввиду этого на рисунках 11, 15, 18 и 19, где длины аминокислотных последовательностей бактериоцинов сравнительно не велики (от 39 до 57 а.о.), мы наблюдаем структуры с визуально меньшим разрешением, а в случае остальных четырех последовательностей – с большим количеством а.о. – мы можем видеть более «сглаженную» структуру. Такой эффект не отражается на точности расчетов, а возникает за счет того, что по вертикали отдельный элемент изображения соответствует одной аминокислоте, линейная же длина осей уравнена.
Более темные области соответствуют центрам областей длины а с максимальным уровнем координации между информацией единицы в последовательности белка. На рисунке показана иерархически организованная структура.
2.5 ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ ПОЛИСАХАРИДОВ (ПЕКТИНА) ИЗ ПРОДУКТОВ РАСТИТЕЛЬНОГО СЫРЬЯ
Пектииновые вещества, или пектины – полисахариды, образованные остатками, главным образом, галактуроновой кислоты. Присутствуют во всех высших растениях, особенно во фруктах, и в некоторых водных растениях, примером является взморник морской. Пектины, являясь структурным элементом растительных тканей, способствуют поддержанию в них тургора, повышают засухоустойчивость растений, устойчивость овощей и фруктов при хранении. Используются в пищевой промышленности – в качестве структурообразователей (гелеобразователей), загустителей, а также в медицинской и фармацевтической промышленности – в качестве физиологически активных веществ, также в виде модифицированного пектина, с полезными для организма человека свойствами. В промышленных масштабах пектиновые вещества получают
в основном из яблочных и цитрусовых выжимок, жома сахарной свеклы, корзинок подсолнечника, тыквы. Пектины являются энтеросорбентами.
Пектин для применения в пищевой и фармацевтической промышленности – очищенный полисахарид – получают кислотной экстракцией из цитрусовых (лайм, лимон, апельсин, грейпфрут), яблочных выжимок, жома сахарной свеклы, морской травы, или из корзинок подсолнечника. Технологическая схема получения пектина предусматривает его экстрагирование из исходного сырья, очистку, осаждение органическими растворителями, сушку, измельчение и стандартизацию. Последний представляет собой процесс модификации свойств пектина, достигаемой физическими и/или химическими способами, с целью приведения их
в соответствие с технологическими и рецептурными требованиями производства различных групп пищевых и непищевых продуктов. Пектин является гелеобразователем, стабилизатором, загустителем, влагоудерживающим агентом, осветлителем, веществом, облегчающим фильтрование, и средством для капсулирования, зарегистрирован в качестве пищевой добавки E440.
В пищевой промышленности пектин используют в производстве начинок для конфет, производстве фруктовых начинок, кондитерских желейных и пастильных изделий (например, зефир, пастила, мармелад), молочных продуктов, десертов, мороженого, спредов, майонеза, кетчупа, сокосодержащих напитков. В фармацевтической и медицинской промышленности его применяют, например, для капсулирования лекарств.
Важное место в увеличении выпуска пектиносодержащих пищевых изделий в России принадлежит яблочному пектину. Пектин из яблочных выжимок составляет 3035 % мирового объема пектиновых веществ и выпускается
в США, Великобритании, Дании, Италии, Германии, Австрии, Болгарии, Польше и Венгрии.
В свежих яблоках протопектин преобладает перед растворимым пектином и составляет 52,3–97,0 % общего количества. Содержание пектина в яблоках невелико – всего 0,95 0,01 %, в том числе нерастворимого – 0,80 0,01 %,
а растворимого – 0,16 0,01 %. При этом наибольшее количество пектиновых веществ сосредоточено в кожице и в семенных камерах.
Схематично строение яблока представлено на рисунке 20.
Рисунок 20 – Схема строения плода яблони:
А зона эпидермиса; В внешняя паренхима; С паренхима семенной камеры; Д семена
Соотношение объемов зоны эпидермиса (кожицы), паренхимы внешней (мякоти) и семенной камеры в зависимости от сорта яблока составляет примерно 25 : 56 : 19.
Внешняя паренхима (В) является структурной основной зоной плода и составляет более, чем 80 % к сухой массе (рисунок 21).
Химический состав зон плода яблони различен, что подтверждают данные, приведенные в таблице 23.
Содержание пектиновых веществ распределяется по зонам следующим образом: А – 4,7–5,2 % к массе сухих веществ; В – 1,8–1,9 %; С и Д – 5,1–5,3 %. При этом наибольшая доля протопектина содержится в кожице, а водорастворимого пектина – в мякоти. Моносахаридный состав пектиновых полисахаридов различных зон яблока также отличается (таблица 24). При этом существенно отличается фракционный состав пектиновых веществ.
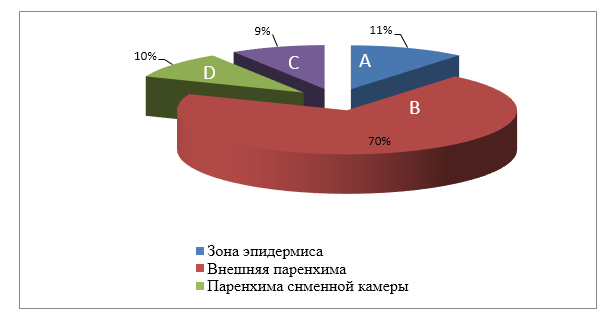
С
Рисунок 21 – Распределение основных структурных зон в плодах яблони
Таблица 23 Химический состав различных зон яблока, мг/г
| Зоны | Углеводный состав спирторастворимых веществ | Белок | Липиды | |||||||
| Rha | Ara | Xyl | Man | Gal | Glc | GalA | Общее | |||
| А | 15 | 47 | 34 | 19 | 54 | 154 | 265 | 588 | 57 | 290 |
| В | 13 | 83 | 74 | 24 | 98 | 296 | 287 | 875 | 27 | 55 |
| С | 11 | 48 | 88 | 23 | 48 | 301 | 283 | 802 | 44 | 62 |
| Д | 13 | 32 | 135 | 28 | 27 | 344 | 253 | 832 | 55 | 38 |
Таблица 24 Моносахаридный состав пектиновых полисахаридов в различных зонах плода яблони
| Фракция | Зоны плода |
Углеводный состав (mole %) | ||||||||
| Rha | Ara | Xyl | Man | Gal | Glc | GalA | Степень
метокси- лиров |
Общее, мг/г |
||
| Раство-
римый пектин |
А | 1,5 | 9,6 | 0,8 | 0,5 | 6,3 | 1,3 | 80,0 | 70 | 582 |
| В | 1,8 | 22,7 | 1,0 | 0,2 | 11,1 | 0,4 | 62,8 | 71 | 722 | |
| С | 1,9 | 5,9 | 0,6 | следы | 5,4 | 0,2 | 86,0 | 62 | 516 | |
| Д | 2,4 | 7.4 | 1,3 | 1,0 | 5,8 | 1,3 | 80,8 | 65 | 428 | |
| Прото-
пектин |
А | 4,9 | 9,7 | 2,9 | 0,7 | 8,2 | 1,8 | 71,8 | 52 | 650 |
| В | 5,7 | 9,0 | 4,5 | 0,4 | 13,2 | 2,9 | 64,3 | 41 | 795 | |
| С | 6,2 | 10,7 | 4,6 | 0,4 | 8,4 | 2,3 | 67,4 | 55 | 759 | |
| Д | 7,1 | 8,4 | 4,2 | 0,8 | 7,3 | 1,9 | 70,3 | 49 | 692 | |
Содержание пектиновых веществ в яблоках зависит также от сорта и района произрастания. Так, в 122 образцах яблок Белоруссии содержание пектина составило от 0,42 до 0,79 %, в 129 – от 0,8 до 0,99 %, в 104 – от 1,0 до 1,5 %;
в 13 – от 1,51 до 1,8 % и в 1 – 2,52 %.
В яблоках среднего Урала в 30 образцах содержание пектина составило до 1,0 %, в 130 – до 1,3 %, в 70 – до 1,9 %. Наибольшее количество пектина содержится в яблоках позднеспелых сортов.
При хранении яблок соотношение фракций пектиновых веществ изменяется. Общее содержание пектина, как правило, уменьшается. Так, в яблоках сорта Джонатан в начале хранения содержится 0,78 %, сорта Голден Делишес – 0,80 % пектиновых веществ, а в конце хранения остается 0,54 и 0,59 % соответственно.
Для получения пектина применяют в основном выжимки среднеспелых
и позднеспелых сортов яблок. Однако установлено, что и раннеспелые сорта Кубани по содержанию пектиновых веществ в период их биологической зрелости (2,6–2,7 %) вполне пригодны для получения из них пектина, хотя полученный пектин уступает по чистоте пектину из средне- и позднеспелых сортов (40,1 против 69,7 %).
По степени метоксилирования, количеству свободных и связанных карбоксильных групп, молекулярной массе пектин ранне- и позднеспелых сортов яблок не различается.
В зависимости от физико-химических показателей яблочного вторичного сырья выжимок и типа получаемого пектина в каждом климатическом районе существуют особенности технологии их переработки.
Технология получения яблочного пектина включает в себя следующие технологические процессы:
Сушеные яблочные выжимки перед процессом гидролиза–экстрагирования пектиновых веществ промывают трижды водой температурой 3035 С. При получении пектина с низкой скоростью студнеобразования промывку ведут водой более высокой температуры (температура смеси 5560 С). Гидролиз–экстрагирование пектина осуществляют водным раствором азотной кислоты при рН 1,5–2,0 при температуре гидролизной смеси – 7080 С, соотношении сырья и экстрагента – 1 : (1012) в течение 3,0–3,5 ч в экстракторе периодического действия. По истечении времени процесса на прессах отделяют
А-экстракт. Отпрессованные яблочные выжимки снова загружают в экстрактор, заливают водой температурой 4550 С в соотношении 1 : (1214). Повторное экстрагирование пектиновых веществ ведут в течение 1,52 ч, получая
В-экстракт, который отделяют прессованием суспензии. Влажность отпрессованных выжимок должна быть не более 80 %, остаточное содержание в них пектина – 0,8–1,0 %.
А-экстракт и В-экстракт смешивают и отстаивают 24 ч для отделения большей части механических примесей. Среднее содержание сухих веществ
в экстракте – 1,0–1,2 %, в том числе пектиновых веществ – 0,3–0,4 %. Отстоявшийся экстракт сепарируют и фильтруют на кизельгуровом фильтре. Концентрирование экстракта проводят в двухкорпусных вакуум-выпарных установках непрерывного действия. Температура продукта в первом корпусе не должна превышать 7075 С, во втором – 45 С. Содержание сухих веществ в концентрате составляет 67 %, концентрация спиртоосаждаемых пектиновых веществ – 2,5–3,5 %, рН концентрата – 1,7–2,2.
Дальнейшее концентрирование до большего содержания сухих веществ считается нецелесообразным, так как повышаются вязкость и кислотность экстракта. Полученный концентрат охлаждают до 25 С, используя для этого пластинчатый теплообменник. Осаждение пектиновых веществ осуществляют этиловым техническим или ректификованным спиртом, крепостью 9095 %. Объемное количество спирта на один объем пектинового концентрата составляет три части. Во избежание выпадения вместе с пектином минеральных примесей осаждение пектиновых веществ ведут при рН 1,7–1,9. Если значение рН концентрата выше, вводят спирт, подкисленный азотной кислотой.
Получаемая пектиноспиртовая суспензия непрерывно поступает на разделение в центрифугу типа ОГШ-353К-09. После центрифугирования пектиновый коагулят с влажностью 7075 % непрерывно ссыпается в промыватель, в который непрерывно подается спирт с крепостью 70 %об. в соотношении 1 : 8. Суспензия первой промывки поступает самотеком на разделение в центрифугу такого же типа. Сырой пектин снова ссыпается в промыватель, где осуществляется вторая промывка спиртом крепостью 9095 %об. в соотношении 1 : 8. Суспензия второй промывки также поступает в центрифугу для отделения пектина. Очищенный пектин подают на сушку.
Сушку осуществляют в барабанной вакуум-сушилке при 5560 С, оборудованной устройствами для удаления конденсированного спирта без остановки процесса, улавливания паров спирта и пектиновой пыли. Продолжительность сушки 23 ч. Сухой пектин измельчают, сушат и направляют на купажирование.
В научно-исследовательской работе использовали пищевой пектин (в соответствии с требованиями нормативного документа «ГОСТ 29186-91. Пектин. Технические условия») – который должен представлять собой порошок тонкого помола без посторонних примесей, однако, допускается наличие волокнистой фракции пектина в виде хлопьев; слабокислый, без запаха, цвет ‒ от светло-серого до кремового, массовая доля влаги – не более 10 %, степень этерификации – от 60 до 70 % в зависимости от типа, студнеобразующая способность – 170 или 200 (не менее, в зависимости от сорта), массовая доля нитратов в расчете на ион NO – не более 0,18 %, массовая доля частиц волокнистой фракции размером более 0,5 мм должна быть не более 20 % и посторонние примеси, видимые невооруженным глазом, должны отсутствовать.
2.6 ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ ПОЛИСАХАРИДОВ (АЛЬГИНАТОВ) ИЗ ВОДОРОСЛЕЙ
Водоросли составляет подавляющую часть фитобентоса Мирового океана, являясь уникальным сырьем для получения ценных пищевых, технических
и медицинских продуктов. Морские водоросли обладают кумулятивной способностью к накоплению разнообразного комплекса микроэлементов, в частности йода, в количествах, значительно превышающих их содержание в окружающей среде.
Уникальной особенностью водорослей является их способность синтезировать и накапливать комплекс полиуроновых кислот, известных под названием «альгиновая кислота», которая в виде альгинатов – солей кальция, магния, натрия – содержится в макрофитах, принадлежащих к родам Macrocystis, Laminaria, Fucus (Наумов И. А. [и др.], 2014).
Альгиновая кислота – полимер уроновых (гулуроновой и маннуровой кислот, соединенных по 1–4 глюкозидным связям) различной степени полимеризации (СП), в результате чего ее молекулярный вес и реологические свойства значительно варьируют. Альгиновая кислота и ее соли способны к 200–300-кратному поглощению воды, образуя гели, для которых характерна высокая кислотоустойчивость. В пищевой промышленности они используются в качестве эмульгаторов, стабилизаторов, желирующих и влагоудерживающих компонентов. В тканях водорослей альгиновые кислоты находятся в форме калиевых, натриевых или кальциевых солей, входящих в состав клеточных стенок, локализованных в межклеточных пространствах слизевых каналов. Водоросли используются в качестве источника альгинатов с целью получения из них неочищенных биогелей, что является альтернативой и позволяет решать многие проблемы, связанные с комплексным использованием сырья и обеспечением населения альгинатсодержащими продуктами (Hecht H., Srebnik S., 2016).
Одной из солей альгиновой кислоты, нашедшей широчайшее использование, является альгинат натрия. Это вещество представляет собой натриевую соль альгиновых кислот, хорошо растворимую в воде.
Альгинат натрия – полисахарид, построенный из единиц D-маннуроновой кислоты, соединенных β–(1,4)–глюкозидными связями. Молекулярный вес альгината натрия колеблется от 50000 до 200000 и зависит от степени полимеризации, которая может достигать 750.
Выпускается для пищевых и технических целей. Образцы отличаются друг от друга чистотой и величиной основных показателей. Альгинат натрия хорошо растворим в воде. Альгинаты биосовместимы, биодеградируемы и относительно недороги в производстве, хорошо растворяются в воде и обладают высокой водоудерживающей способностью, легко образуют гидрогели. Благодаря всему этому альгинаты нашли широкое применение в качестве загустителей, гелеобразователей и стабилизаторов в медицине и фармацевтике, а также в текстильной индустрии. Но особенно широко применение альгинатов в сфере производства продуктов питания, в частности – соусов, паст, желирующих заливок, крем-супов, мороженого и т. д.
Технология получения альгината натрия включает в себя следующие этапы:
Измельченные до размера 3‒5 мм или воздушно-сушеные водоросли, предварительно замоченные и промытые в воде, смешивают с 1‒2%-м раствором поваренной соли в соотношении 1 : 10‒1 : 15, обрабатывают в анодной камере диафрагменного электролизера, снабженного рубашкой с подогревом, до достижения рН ≤ 2,0 и выдерживают при температуре 40‒50 °С
в течение 0,5‒1 ч.
В другом варианте, после электрохимической обработки, т. е. после достижения рН ≤ 2,0, при комнатной температуре в анодной камере элекролизера без тепловой рубашки смесь водорослей с раствором электролита сливают в емкость (реактор с подогревом и мешалкой) и выдерживают при температуре 40‒50 °С при постоянном перемешивании в течение 2 ч.
Отделенные от раствора электролита и промытые водоросли заливают 1‒2 % раствором NaCl в соотношении 1 : 30‒1 : 50 и обрабатывают в катодной камере электролизера, снабженного рубашкой для подогрева, при рН ≥ 12,0 и температуре 60 °С при постоянном перемешивании в течение 0,5‒1 ч (альгиновая варка) до получения однородной вязкой массы, т. е. до полного экстрагирования альгинатов из обрабатываемого сырья, или, в другом варианте, после достижения рН ≥ 12,0 в катодной камере электролизера без тепловой рубашки смесь переводят в реактор с мешалкой и подогревом и выдерживают в нем при температуре 50‒60 °С и постоянном перемешивании в течение 2‒3 ч. Далее смесь фильтруют и охлаждают до 18 °С. Фильтрат помещают в анодную камеру электролизера и обрабатывают при постоянном перемешивании до достижения рН ≤ 2,0 и полного осаждения альгиновой кислоты (перевод в нерастворимую форму), которую затем промывают холодной водой. Промытую альгиновую кислоту смешивают с горячей водой при температуре 80‒90 °С в соотношении 1 : 2 и выдерживают 1‒2 ч для коагуляции. Воду сливают и дают альгиновой кислоте обтечь в течение 30 мин. Осадок альгиновой кислоты сушат и расфасовывают. Полученная светлая альгиновая кислота представляет собой готовый продукт или может служить полуфабрикатом для получения альгината натрия.
Для получения альгината натрия альгиновую кислоту смешивают с 0,5‒1,0 % раствором NaCl в соотношении 1 : 10 и обрабатывают в катодной камере электролизера при слабощелочной реакции среды (рН не выше 8,0)
в течение 0,5 ч. Раствор альгината натрия направляется в бак-накопитель,
а затем на сушку, дробление, расфасовку и упаковку. Образующиеся стоки после анодной и катодной обработки взаимно нейтрализуются.
В научно-исследовательской работе использовали пищевой альгинат натрия (в соответствии с требованиями нормативного документа ТУ 15-544-83 «Альгинат натрия пищевой. Технические условия») – который должен представлять собой сухой порошок или небольшие тонкие пластинки, его окраска колеблется в зависимости от сырья – от кремовой (из ламинарных водорослей) до коричневой (из фукусов). Массовая доля влаги в продукте не должна быть выше 16 %; массовая доля золы (в пересчете на сухое вещество) – не выше 26 %; количество веществ, не растворимых в кипящей воде – не выше 0,3 %; содержание альгиновой кислоты – не менее 70 % (в пересчете на сухое вещество); кинематическая вязкость 1 %-го раствора альгината натрия (в пересчете на сухое вещество) при температуре 20 °С не должна быть ниже 30 сСт.
2.7 ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ ПОЛИСАХАРИДОВ (КСАНТАНА) МИКРОБНОГО ПРОИСХОЖДЕНИЯ
Ксантановая камедь (ксантан) природное химическое соединение (C35H49O29)n, пищевая добавка Е415, относится к группе стабилизаторов.
По химической природе ксантановая камедь представляет собой полисахарид, полученный путем ферментации с использованием бактерии Xanthomonas campestris. В жизненном цикле бактерий служит им защитой от вирусов и пересыхания, поэтому может использоваться в косметических средствах для увлажнения кожи. Производство ксантана основано на аэробном брожении в водном растворе углеводов, источника азота и др., после чего среду пастеризуют
и осаждают спиртом или очищают методом микрофильтрации.
Свойства ксантана регулируют, изменяя условия жизни бактерий. Главная цепь полимера идентична молекуле целлюлозы. Ответвления представляют собой остатки молекул глюкозы, маннозы, глюкуроновой кислоты, а также пировинограднокислые (пируватные) и ацетильные группы. Число пируватных групп определяет вязкость водных растворов ксантана. Для пищевых целей кислотные группы нейтрализуют, переводя ксантан в калиевые, натриевые или кальциевые соли.
Впервые ксантан был получен в конце 1950-х гг. в Северной Региональной научно-исследовательской лаборатории (NRRL) Министерства сельского хозяйства США группой исследователей во главе с Розалиндой Джинсе, которая вместе с другими исследователями, внесла заметный вклад в исследование структуры этого полисахарида.
Экзополисахариды фитопатогенных бактерий рода Xanthomonas являются гетерополисахаридами. Моносахаридный состав их сходен. В большинстве случаев они содержат D-глюкозу, D-маннозу, D-глюкуроновую кислоту. При сходном качественном составе, существуют значительные отличия в молярном соотношении даже у штаммов Xanthomonas одного вида – у некоторых преобладает глюкоза, у других соотношение глюкозы и маннозы одинаково или сдвинуто в сторону маннозы. Углеводный состав ксантана сложно изучить из-за высокой устойчивости целлюлозной основы к гидролизу.
Основная цепь ксантана состоит из (1→4)глюкопиранозы. Каждый второй остаток глюкопиранозы имеет трисахаридную боковую цепь, которая прикрепляется к остатку глюкопиранозы основной цепи. Боковая цепь содержит два остатка маннозы и остаток глюкуроновой кислоты. Одна молекула маннопиранозы соединена с остатком глюкопиранозы основной цепи связью (1→3),
а с глюкуроновой кислотой β 1→2 связью. Конечный β–маннопиранозный остаток связан с остатком β-глюкуроновой кислоты связью 1→4, которая,
в свою очередь, (1→2) связью прикреплена к неконечному остатку
α-маннопиранозы. Одна молекула α-маннопиранозы ацетилирована в положении О – 6, маннопираноза, находящаяся в конце боковой цепи включает пировиноградную кислоты. Химический состав ксантана может варьироваться в зависимости от способа получения биополимера.
Химическое строение молекул ксантана обеспечивает ему многообразие функционально-технологических свойств: высокую растворимость в холодной и горячей воде; стабильность в растворах при низких и высоких температурах
и широком диапазоне рН от 1 до 13, к действию ферментов; к предотвращению синерезиса (отделение влаги) в продукте и др.
В пищевой промышленности ксантан используют при производстве молочных пудингов, сметаны, йогуртов, мороженого, майонеза и других пищевых продуктов (Липатов Н. Н., 1987; Губанова Т. С., 2001; Горощенко Г., 2003).
В косметической промышленности ксантан применяют при изготовлении солнцезащитных лосьонов и кремов, в том числе для автозагара; очищающих средств и кремов для лица, тела; краски для волос; шампуней и гелей для душа; зубных паст.
В фармацевтической индустрии ксантан находит применение при производстве пероральных лекарственных препаратов (капсулы, порошки) и препаратов местного действия (мази, гели, линименты). При использовании 5 % этого вещества изменяются реологические свойства коллоидной системы, что положительно влияет на структуру производимых продуктов. Последние годы получило развитие производство микробных экзополисахаридов в сравнении
с получением полисахаридов из высших организмов. К числу таких веществ принадлежат полисахариды, использующиеся в медицинской и пищевой отраслях промышленности (Шацкая Н. Г., 1994; Забодалова П. А., 2000; Wessels S., 2004). Это связано с тем, что микробные полисахариды имеют более разнообразные функциональные свойства, легко воспроизводимы и безопасны, получаются независимо от времени года с наименьшими затратами (Гвоздяк Р. И., 1989).
Ряд авторов отмечают, что ксантан является иммуностимуляторам
и проявляет лечебно-профилактические свойства при язвенной болезни, гипертонии, атеросклерозе, онкологических заболеваниях (Шендеров Б. А., 1998; Мартынов А. В., 1999).
Основными физическими свойствами полисахаридов являются способность к набуханию и растворению в холодной или горячей воде с образованием гелей. На эти свойства в зависимости от вида полисахарида влияют следующие факторы: температура, активная pH среды, структура и его состав, происхождение, а также степень очистки. Применение полисахарида в пищевой промышленности в качестве структурообразователей определяется их технологическими свойствами.
В последние годы, в связи с увеличивающимся объемом производства кисломолочных продуктов с длительным сроком хранения, повысился интерес к закваскам, образующим вязкие сгустки, которые обладают влагоудерживающей способностью. Было установлено, что такие закваски при сквашивании молока продуцируют экзополисахариды, которые являются естественными загустителями, заменяющими стабилизаторы растительного и животного происхождения.
Особенно важным является использование штаммов продуцирующих ксантан для улучшения реологических свойств и функциональных характеристик кисломолочных продуктов в странах, где запрещено использование стабилизаторов. Эта особенность на сегодняшний день находится в центре внимания среди производителей продуктов питания.
На основании всего вышеизложенного, можно выделить основные функции ксантана, которые имеют важное значение в пищевой, косметологической и фармацевтической промышленностях:
- стабилизирует эмульсии и суспензии в широком диапазоне температур;
- при механическом воздействии (при перемешивании, перекачке и наполнении тары) вязкость продукта, стабилизированного ксантаном, понижается с последующим восстановлением;
- способствует пенообразованию и стабилизирует пену;
- образует прозрачный раствор, пригодный для загущения прозрачных продуктов;
- легко диспергируется.
Технология получения ксантана включает в себя следующие этапы:
В состав ферментационной среды для культивирования продуцента (Xanthomonas campestris) ксантана входят источник сахаров (упаренный гидролизат древесного сырья – УГДС), кукурузный экстракт (ростовой фактор), аммоний сернокислый, калий фосфорнокислый однозамещенный, магний сернокислый и вода.
Основой источника сахаров является отход – древесная щепа. Размер исходных частиц щепы варьирует в широком пределе и зависит от вида древесины. Гидролиз щепы может быть как кислотным, так и ферментативным. В любом варианте гидролиз проводят известным образом.
Выращивание посевного материала проводят на скошенной агаризованной среде. В качестве среды для твердофазного культивирования используют агаризованный 2 % агара бульон МПА с добавлением 2 % стерильной глюкозы. Полученную среду в асептических условиях разливают в стерильные пробирки
с ватными пробками. Укладывают пробирки на валик для получения скошенной твердой среды. После остывания и затвердения агаризованной среды
в пробирках, их помещают в термостат при температуре 37 °C на 5 сут для проращивания.
Через 5 сут отбирают визуально стерильные пробирки со скошенной агаризованной средой и производят в асептических условиях посев культуры продуцента ксантана микробиологической петлей с пробирки с культурой, проверенной на отсутствие контаминантов визуально и микроскопированием, хранящейся в холодильнике не более 20 сут.
Пробирки после посева помещают в термостат при температуре 2728 °C на 24 ч. Посевная среда для выращивания в колбах имеет следующий состав, г/л: УГДС (по глюкозе) 10,0; кукурузный экстракт 15,0; аммоний сернокислый 1,0; калий фосфорнокислый однозамещенный 1,5; магний сернокислый пятиводный 0,7; водопроводная вода остальное.
В асептических условиях посевную среду по 10 мл заливают в пробирки
с культурой, полученной ранее, и производят смыв культуры с поверхности агаризованной среды стерильной пипеткой. Среду с культурой из пробирок асептически переносят в посевную среду. Перемешивают и разливают по стерильным колбам емкостью 250 мл по 30 мл в каждую колбу. Приготовленные колбы устанавливают на качалке. Включают перемешивание на 380400 об./мин и устанавливают температуру культивирования 2628 °C.
В установленном режиме колбы культивируют в течение 24 ч. Затем снимают их с качалки и в асептических условиях сливают посевной материал из каждой колбы в стерильную колбу с нижним тубусом, шлангом и зажимом на нем. Из этой колбы отбирают пробу усредненного посевного материала из колб и осуществляют проверку посевного материала на отсутствие контаминантов микроскопированием и высевом на твердую питательную среду.
При культивировании продуцента ксантана в биореактор, в котором находится 8 л стерильной УГДС, вносят ранее полученный посевной материал, а также ранее подготовленные стерильные растворы солей и ростового фактора. Включают подачу стерильного воздуха в биореактор. Процесс проводят при термостатировании и перемешивании содержимого биореактора.
В процессе культивирования из биореактора периодически отбирают пробу на отсутствие контаминантов, оптическую плотность и содержание редуцирующих веществ (РВ, по глюкозе). По показаниям контрольного pH-метра корректируют значение pH в начальной ферментационной среде. При соблюдении всех вышеперечисленных условий, в асептических условиях присоединяют к биореактору колбу с 2 л посевного материала и переносят содержимое колбы в биореактор. Затем культивирование продуцента ксантана осуществляют в биореакторе.
В течение всего процесса культивирования из биореактора периодически отбирают пробу культуральной жидкости для контроля на отсутствие контаминантов, определения концентрации биомассы и ксантана и концентрации остаточных РВ. При необходимости проводят корректировки состава питательной среды.
Окончание процесса культивирования определяется по результатам анализа проб из биореактора на содержание РВ и показаниям датчика измерения парциального давления растворенного кислорода. По окончании процесса культивирования культуральную жидкость сливают из биореактора через нижний штуцер в пластиковые канистры и культуральная жидкость поступает на стадию выделения ксантана.
Культуральную жидкость обрабатывают этиловым спиртом при перемешивании, отделяют спиртовую фракцию, из которой любым известным способом (предпочтительно, выпариванием спирта) выделяют ксантан.
В научно-исследовательской работе использовали ксантан пищевой
(в соответствии с требованиями нормативного документа ГОСТ 333332015 «Добавки пищевые. Камедь ксантановая Е415. Технические условия»), который должен представлять собой белый, кремово-белый или серовато-белый сыпучий порошок, без запаха, растворим в дистиллированной воде и нерастворим в этиловом спирте. По физико-химическим показателям ксантан при проведении теста на гелеобразование должен давать положительный результат, массовая доля основного вещества в высушенной ксантановой камеди 91,0108,0 %, что соответствует массовой доли диоксида углерода (СО2) 4,25,0 %; массовая доля потерь при высушивании – не более 15,0 %; массовая доля потерь при прокаливании – не более 16,0 %; массовая доля пировиноградной кислоты – не менее 1,5 %; массовая доля азота – не более1,5 %; массовая доля этилового или изопропилового спирта, или суммарно – не более 0,05 %.
2.8 ВЛИЯНИЕ ПОЛИСАХАРИДНЫХ КОМПОНЕНТОВ РАЗЛИЧНОГО ПРОИСХОЖДЕНИЯ НА ВЫЖИВАЕМОСТЬ МОЛОЧНОКИСЛЫХ БАКТЕРИЙ
Немаловажным показателем пробиотических свойств молочнокислой флоры является их способность колонизировать желудочно-кишечный тракт,
а также их резистентность к биологическим жидкостям организма-хозяина (Андрейчин М. А., 1980; Петров Л. Н., 2002; Буряко И. А. [и др.], 2004). Последний показатель визуализировали по наличию или отсутствию роста изучаемой микрофлоры в жидкой питательной среде, содержащей натуральный желудочный сок лошади (препарат «Эквин»), желчь крупного рогатого скота медицинская консервированная (20 %, 30 %, 40 %), фенол (0,4 %) и NaCl (2 %, 4 %) (Лабинская А. С., 1978; МУ 2.3.2.2789-10). Результаты по влиянию агрессивных сред на выживаемость лактобактерий без применения полисахаридных компонентов представлены в таблице 25.
Таблица 25 – Устойчивость лактобацилл к агресивным средам
без полисахаридов
| Штамм | Рост в питательной среде, содержащей NaCl | Рост в питательной среде, содержащей желчь |
Рост в питательной среде, содержащей фенол (0,4 %) | Рост в питательной среде, содержащей желудочный сок | |||
| 2 % | 4 % | 20 % | 30 % | 40 % | |||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| Примечание: «+» – хороший рост; «±» – слабый рост. | |||||||
Результаты исследований показали, что культуры изучаемых лактобактерий проявили хороший рост только на среде с содержанием 2,0 и 4,0 % хлорида натрия, а также при наличии в среде желчии – в концентрации 20 %. Незначительный рост дала культура Lactobacillus salivarius PC20b на питательной среде, содержащей 30 % желчи. В остальных случаях рост исследуемых культур
не проявился.
Результаты влияния различных доз микробных полисахаридов (ксантана) на выживаемость лактобацилл, представлены в таблице 26.
Таблица 26 – Устойчивость лактобацилл к агресивным средам с добавлением ксантана
| Штамм | Рост в питательной среде, содержащей NaCl | Рост в питательной среде, содержащей желчь |
Рост в питательной среде, содержащей фенол (0,4 %) | Рост в питательной среде, содержащей желудочный сок | |||
| 2 % | 4 % | 20 % | 30 % | 40 % | |||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 0,1 % ксантана | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| 0,2 % ксантана | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| 0,3 % ксантана | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Продолжение таблицы 26 | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| 0,4 % ксантана | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| 0,5 % ксантана | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| Примечание: «+» – хороший рост; «±» – слабый рост. | |||||||
Проведенные результаты исследований показали, что использование в составе питательных сред ксантана в изучаемых концентрациях (0,1‒0,5 %) не оказало влияния на жизнеспособность изучаемых лактобактерий при действии на них агресивных растворов. Из таблиц 26 видно, что культуры изучаемых лактобактерий проявили хороший рост, как и в случае отсутствия полисахаридных компонентов, только на среде с содержанием 2,0 и 4,0 % хлорида натрия,
а также при наличии в среде желчи в концентрации 20 %. В остальных вариантах рост изучаемых лактобацилл не проявился или был незначительным.
Результаты влияния различных доз полисахаридов из водорослей (альгината натрия) на выживаемость лактобацилл представлены в таблице 27.
Серия проведенных экспериментов с использованием в составе питательных сред различных доз альгината натрия (таблица 27) показала, что в дозе полисахарида 0,3; 0,4 и 0,5 % наблюдается хороший рост лактобацилл не зависимо от действия на них изучаемых агрессивных растворов.
Таблица 27 – Устойчивость лактобацилл к агресивным средам с добавлением альгината натрия
| Штамм | Рост в питательной среде, содержащей NaCl | Рост в питательной среде, содержащей желчь |
Рост в питательной среде, содержащей фенол (0,4 %) | Рост в питательной среде, содержащей желудочный сок | |||
| 2 % | 4 % | 20 % | 30 % | 40 % | |||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 0,1 % альгинат натрия | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | − | ± | − |
| Lactobacillus curvatus PC723_5 | + | + | + | + | − | ± | − |
| Lactobacillus sp. PC18A | + | + | + | + | − | ± | − |
| 0,2 % альгинат натрия | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | ± | + | ± |
| Lactobacillus curvatus PC723_5 | + | + | + | + | ± | + | ± |
| Lactobacillus sp. PC18A | + | + | + | + | ± | + | ± |
| 0,3 % альгинат натрия | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | + | + | + |
| Lactobacillus curvatus PC723_5 | + | + | + | + | + | + | + |
| Lactobacillus sp. PC18A | + | + | + | + | + | + | + |
| 0,4 % альгинат натрия | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | + | + | + |
| Lactobacillus curvatus PC723_5 | + | + | + | + | + | + | + |
| Lactobacillus sp. PC18A | + | + | + | + | + | + | + |
| 0,5 % альгинат натрия | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | + | + | + |
| Продолжение таблицы 27 | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Lactobacillus curvatus PC723_5 | + | + | + | + | + | + | + |
| Lactobacillus sp. PC18A | + | + | + | + | + | + | + |
| Примечание: «+» – хороший рост; «±» – слабый рост. | |||||||
Результаты влияния дозличных доз полисахаридов растительного происхождения (пектина) на устойчивость лактобактерий к агрессивным средам представлены в таблице 28.
Таблица 28 – Устойчивость лактобацилл к агресивным средам с добавлением пектина
| Штамм | Рост в питательной среде, содержащей NaCl | Рост в питательной среде, содержащей желчь |
Рост в питательной среде, содержащей фенол (0,4 %) | Рост в питательной среде, содержащей желудочный сок | |||
| 2 % | 4 % | 20 % | 30 % | 40 % | |||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 0,1 % пектина | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | − | − |
| Lactobacillus curvatus PC723_5 | + | + | + | − | − | − | − |
| Lactobacillus sp. PC18A | + | + | + | − | − | − | − |
| 0,2 % пектина | |||||||
| Lactobacillus salivarius PC20b | + | + | + | ± | − | ± | − |
| Lactobacillus curvatus PC723_5 | + | + | + | ± | − | ± | − |
| Lactobacillus sp. PC18A | + | + | + | ± | − | ± | − |
| 0,3 % пектина | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | − | ± | − |
| Lactobacillus curvatus PC723_5 | + | + | + | + | − | ± | − |
| Продолжение таблицы 28 | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| Lactobacillus sp. PC18A | + | + | + | + | − | ± | − |
| 0,4 % пектина | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | + | + | ± |
| Lactobacillus curvatus PC723_5 | + | + | + | + | + | + | ± |
| Lactobacillus sp. PC18A | + | + | + | + | + | + | ± |
| 0,5 % пектина | |||||||
| Lactobacillus salivarius PC20b | + | + | + | + | + | + | + |
| Lactobacillus curvatus PC723_5 | + | + | + | + | + | + | + |
| Lactobacillus sp. PC18A | + | + | + | + | + | + | + |
| Примечание: «+» – хороший рост; «±» – слабый рост. | |||||||
Из таблицы видно, что наилучшие результаты исследований проявились
с применением в составе питательной среды, на которой выращивали изучаемые лактобактерии, пектина в экспериментально-максимальной дозе 0,5 %.
Таким образом, проведенные исследования показали, что выделенные из желудочно-кишечного тракта фазана лактобациллы за счет использования полисахаридного компонента способны проявлять резистентность к фенолу, желчи, хлориду натрия и желудочному соку, что позволяет предложить данные культуры микроорганизмов в качестве основных объектов для конструирования биопрепарата с высокой пробиотической активностью, а также рекомендовать для максимальной выживаемости лактобацилл, в состав пробиотического препарата Галлобакт-Ф вносить полисахаридную смесь на основе альгината натрия в дозе 0,3 %.
2.9 ТЕХНОЛОГИЯ ПОЛУЧЕНИЯ ПРОБИОТИКА ГАЛЛОБАКТ-Ф-ПЛЮС
Галлобакт-Ф-Плюс включает в себя ассоциацию трех видов молочнокислых бактерий: Lactobacillus salivarius PC20b, Lactobacillus curvatus PC723_5, Lactobacillus sp. PC18A с добавлением в состав пробиотика полисахарада ‒ альгината натрия. Все штаммы не являются генномодифицированными.
Готовая добавка состоит из жизнеспособных штаммов молочнокислых бактерий и имеет титр в количестве не менее 1,0 × 109 КОЕ/см3 (колониеобразующих единиц) и представляет жидкость со слабым, специфическим для данного продукта запахом, так как содержит вспомогательные вещества – воду, мелассу свекловичную, молоко или молочную сыворотку, а также альгинат натрия.
Технологический процесс производства Галлобакт-Ф-Плюс включает в себя следующие стадии:
- получение чистых культур Lactobacillus salivarius PC20b, Lactobacillus curvatus PC723_5, Lactobacillus sp. PC18A и их хранение;
- размножение чистых культур Lactobacillus salivarius PC20b, Lactobacillus curvatus PC723_5, Lactobacillus sp. PC18A на обезжиренном молоке по отдельности в течение 1–3 сут при температуре 30–35 °С в пробирках в лабораторных условиях;
- приготовление маточной культуры (культурной жидкости) в колбах
и трехлитровых банках на основе данных микроорганизмов; - культивирование штаммов с добавлением полисахаридных компонентов растительного происхождения в ферментере;
- расфасовка препарата;
- хранение готовой продукции.
Используемые культуры микроорганизмов имеют постоянный размер – это небольшие палочки размером от 3–4 мкм и толщиной до 0,5 мкм (рисунки 22–24). Полиморфизм, который очень часто наблюдается у отдельных видов лактобацилл, не выявлен. В процессе жизнедеятельности клетки становятся короче и уменьшаются в размере, что связано с их старением.
Рисунок 22 Микроскопия клеток Lactobacillus salivarius PC20b через 24 ч культивирования
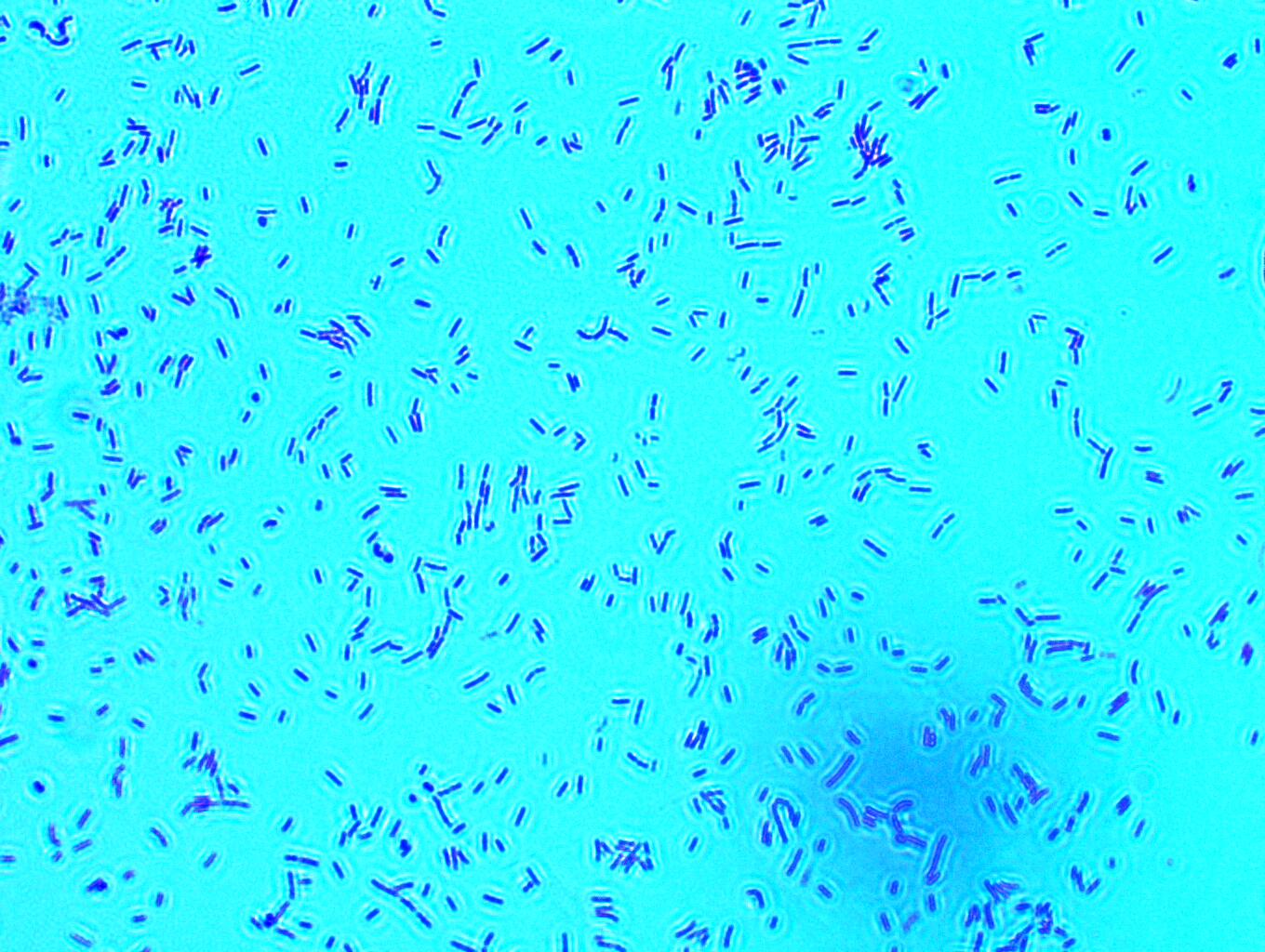
Рисунок 23 Микроскопия клеток Lactobacillus sp. PC18A через 24 ч
культивирования
Рисунок 24 Микроскопия клеток Lactobacillus curvatus PC723_5 через 24 ч культивирования
Размножение штаммов микроорганизмов в пробирках в лабораторных условиях. Культуры лактобактерий засевают в стерильное обезжиренное молоко (обрат) в пробирки. Культивируют 72 ч при температуре 37 ºС.
Культивирование штаммов микроорганизмов в трехлитровых емкостях. Питательная среда для маточной культуры I генерации бактерий готовится тоже на основе обезжиренного молока, разведенного в два раза водопроводной водой.
Маточная культура II генерации бактерий в трехлитровых банках культивируется на обезжиренном молоке (обрате), разведенном в три раза водопроводной водой.
Рост маточной культуры I и II генерации длится 5–7 дней при температуре 35–38 ºС. После этого производят контроль чистоты культур. При хорошем качестве маточной культуры ее передают для засева в ферментеры.
Культивирование используемых штаммов в ферментерах. Производственная среда для бактерий готовится на основе обезжиренного молока или молочной сыворотки (четвертая часть от объема среды), воды водопроводной и 2,0 % мелассы. В качестве компонента, обеспечивающего пребиотические свойства добавки, а также максимальную выживаемость культур лактобактерий в желудочно-кишечном тракте, добавляют 0,3 % смеси полисахарида растительного происхождения (альгинат натрия). В подготовленные питательные среды вносят маточные закваски в количестве 3– % от объема среды. Ферментация культур длится 2−3 сут при температуре 35–38 ºС в анаэробных условиях. Во время ферментации регулярно берутся пробы для контроля происходящего процесса. Контролируется рН препарата, чистота культивируемого штамма. В конце процесса берутся пробы для определения титра. Титр лактобактерий в жидкой форме препарата должен быть не менее 50 млн живых бактериальных клеток в 1 см3.
Получение готового продукта Галлобакт-Ф-Плюс заключается в совместном культивировании молочнокислых микроорганизмов: Lactobacillus salivarius PC20b, Lactobacillus curvatus PC723_5, Lactobacillus sp. PC18A.
Оснащение и подготовка ферментера. Ферментер должен быть оснащен необходимым оборудованием для выполнения следующих технологических операций: культивирование штаммов молочнокислых микроорганизмов; перемешивание; рН-статирование; регистрация значения рН в постоянном режиме; посев через инокуляционную емкость с помощью перистальтического насоса.
Приготовление питательной среды. После полного растворения всех компонентов среды, доводят рН раствора до значения 7,0–7,2, используя 25%-й водный аммиак.
Стерилизация питательной среды. Стерилизацию осуществляют по следующему режиму: нагрев и выдержка при температуре 121 ± 1 С в течение
20 мин для удаления воздуха из ферментера; нагрев до температуры 121 ± 1 С (1,0 атм.) с выдержкой при этой температуре 30 мин и пропаркой всех отводов; охлаждение питательной среды до температуры 37 С, поддерживая избыточное давление в полости ферментера 0,04 ± 0,01 МПа.
В момент стерилизации питательной среды для штаммов молочнокислых микроорганизмов инокуляционная емкость с маточной культурой и емкость
с раствором 25%-го водного аммиака должны быть установлены и присоединены к ферментеру.
После достижения питательной средой температуры 37 С значения рН необходимо установить на уровне 7,0. В случае необходимости микробиолог сообщает оператору и асептически осуществляет контрольный отбор пробы для химического анализа и контроля стерильности.
Инокуляция маточной культуры. Инокуляция осуществляется путем асептической передачи жидкой маточной культуры из инокуляционной емкости
в ферментер с помощью перистальтического насоса. В момент передачи в полости ферментера должно поддерживаться избыточное давление на уровне 0,01−0,02 МПа.
Режим культивирования. Культивирование штаммов молочнокислых микроорганизмов осуществляется в течение 20−22 ч с соблюдением следующих параметров:
- температура – 37–40 С;
- перемешивание – 15–20 об./мин.;
- аэрирование – отсутствует (избыточное давление воздухом 0,04 МПа).
Оператор сообщает микробиологу о необходимости отбора контрольной пробы через 16–18 и 30–32 ч культивирования. Отбор пробы осуществляется через предварительно пропаренный в течение 15 мин пробоотборник.
Завершение ферментации. На основании анализа контрольной пробы микробиолог дает заключение о завершении процесса культивирования. Перед розливом готового продукта в тару хранения оператор сообщает микробиологу
о необходимости отбора проб готовой продукции. В течение часа после расфасовывания готовый продукт должен быть размещен на хранение в холодильной камере либо реализован потребителю.
Контрольные точки.
- стерильность воздуха перед входом в ферментер (при необходимости);
- контроль стерильности и химический анализ питательной среды после стерилизации (при необходимости);
- контрольная проба продукта через 16–18 ч (обязательно);
- контрольная проба готового продукта (обязательно).
Расфасовка препарата. Галлобакт-Ф-Плюс расфасовывают по 2,0; 5,0; 10,0 дм3 в герметичную тару, изготовленную из полимерных материалов по ГОСТ Р 51760. Емкости обрабатывают спиртом. Каждую упаковочную единицу снабжают инструкцией по применению.
Пробиотическую добавку Галлобакт-Ф-Плюс хранят в чистом, защищенном от света помещении при температуре от 2 до 10 °С. Срок годности препарата – 3 мес со дня изготовления. Галлобакт-Ф-Плюс после истечения срока годности не должен применяться.
2.10 ИЗУЧЕНИЕ БИОЛОГИЧЕСКИХ СВОЙСТВ ИССЛЕДУЕМЫХ КУЛЬТУР
В качестве биохимических свойств выделенных лактобацилл проводили определение спектра сбраживаемости сахаров, потребность молочнокислых микроорганизмов в отдельных витаминах, которое оценивали отсутствием роста в питательной среде без пищевого фактора, а также активное кислотообразование исследуемых лактобацилл – титрометрическим методом (Лабинская А. С., 1978).
Сбраживаемость углеводов, является важным физиолого-биохимическим свойством лактобактерий (таблица 29). Как видно из таблицы 29, исследуемые лактобациллы из девяти используемых сахаров не способны были утилизировать только ксилозу. Глюкозу, арабинозу, мальтозу, целлобиозу, фруктозу, лактозу, раффинозу и сахарозу молочнокислые бактерии легко использовали в качестве пищевого фактора.
Таблица 29 – Сбраживание углеводов молочнокислыми бактериями
| Штамм | Сбраживание* | ||||||||
| глюкозы | целлобиозы | фруктозы | лактозы | раффинозы | сахарозы | ксилозы | арабинозы | мальтозы | |
| Lactobacillus salivarius PC20b |
+ | + | + | + | + | + | – | + | + |
| Lactobacillus curvatus PC723_5 |
+ | + | + | + | + | + | – | + | + |
| Lactobacillus sp. PC18A | + | + | + | + | + | + | – | + | + |
| Примечание: «+» – сбраживается; «–» – не сбраживается. | |||||||||
При подборе штамма-пробионта немаловажно знать трофические способности микроорганизма по таким питательным компонентам как витамины, так как чем меньше штамм будет в них нуждаться, тем проще формировать питательную среду для накопления биомассы клеток. Нами использовались такие витамины как тиамин (витамин В1), рибофлавин (витамин В2) и фолиевая кислота (витамин В9). Потребность испытуемых лактобактерий в витаминах представлена в таблице 30.
Таблица 30 – Потребность молочнокислых бактерий в витаминах
| Штамм | Потребность в* | ||
| тиамине | рибофлавине | фолиевой кислоте | |
| Lactobacillus salivarius PC20b |
– | + | – |
| Lactobacillus curvatus PC723_5 |
– | + | – |
| Lactobacillus sp. PC18A | − | + | – |
| Примечание: «+» – требуется для роста; «–» – не требуется для роста. | |||
Результаты исследований показали, что используемые в работе лактобактерии не сильно требовательны к витаминам, в частности, витамину В1 и В9, так как отсутствие их в качестве пищевого фактора не сказалось на росте микроорганизмов. Однако следует отметить, что присутствие в питательной среде рибофлавина является обязательным фактором роста.
Нормируемым показателем специфической активности лактосодержащих пробиотиков является активность кислотообразования, а значит, одним из важных критериев отбора штаммов при разработке биопрепаратов (Кощаев А. Г., 2008) (таблица 31).
Таблица 31 – Активное кислотообразование исследуемых штаммов
| Штамм | Титруемая кислотность, °Т | ||
| 6 ч | 12 ч | 24 ч | |
| Lactobacillus salivarius PC20b |
43,5 ± 1,3 | 124,7 ± 2,4 | 246,7 ± 4,2 |
| Lactobacillus curvatus PC723_5 |
60,4 ± 1,5 | 132,2 ± 2,1 | 267,6 ± 4,6 |
| Lactobacillus sp. PC18A | 47,7 ± 1,4 | 118,5 ± 2,2 | 241,3 ± 4,1 |
Из таблицы 31 видно, что изучаемые штаммы обладали высокими показателями титруемой кислотности. Наивысшая титруемая кислотность наблюдалась у Lactobacillus curvatus PC723_5, которая на конец выращивания микроорганизмов (через 24 ч) составила ‒ 267,6 °Т, в то время как в питательной среде
с Lactobacillus salivarius PC20b и Lactobacillus sp. PC18A данный показатель составил 246,7 и 241,3 °Т, соответственно.
Таким образом, проведенные исследования показали, что выделенные из желудочно-кишечного тракта дикого фазана лактобациллы обладают свойствами, характерными для молочнокислых бактерий – у них наблюдается высокая кислотообразующая способность, они утилизируют широкий спектр сахаров, что дает им преимущества при конкуренции за субстрат. Все эти свойства позволяют предложить данные культуры микроорганизмов в качестве основных объектов для конструирования биопрепарата с высокой пробиотической активностью.
2.11 ИЗУЧЕНИЕ ФЕНОТИПИЧЕСКОГО ПРОФИЛЯ АНТИБИОТИКОРЕЗИСТЕНТНОСТИ ОТОБРАННЫХ МИКРООРГАНИЗМОВ
К важным технологическим и пробиотическим свойствам молочнокислых микроорганизмов относится их устойчивость к антибиотикам, так как
в промышленном животноводстве, в частности птицеводстве, использование данных препаратов является повседневным (Глушанова Н. А., 2003).
Резистентность штаммов определяли к терапевтическим дозам антимикробных веществ диско-диффузионным методом в агар с использованием стандартных дисков (Балтрашевич А. К., Комаровская Т. П., 1982; Воробьёва Л. И., 1989; МУ 2.3.2.2789-10). Результаты чувствительности используемых штаммов к терапевтическим дозам некоторых антибиотиков представлены в таблице 32.
Результаты оценки антибиотикочувствительности отобранных штаммов показали, что ингибирующие рост препараты в различной степени действовали на лактобациллы. При этом хотелось бы отметить, что согласно критериям оценки чувствительности лактобактерий, с учетом диаметра зоны ингибирования их роста все используемые микроорганизмы можно отнести к категории «устойчивые» к действию применяемых антимикробных веществ (Назырова Н. Р., Тимербаева Р. X., Туйгунов М. М., 2006; МУ 2.3.2.2789-10). Только лишь при действии линкомицина и цефалотина на Lactobacillus sp. PC18A,
а также цефалотина – на Lactobacillus salivarius PC20b была выявлена зона задержки роста в пределах 11 мм, что является минимальным значением, чтобы отнести используемые лактокультуры по отношению к данным антимикробным вещестам к категории «промежуточно устойчивые».
Таблица 32 – Антибиотикочувствительность лактобактерий
| Антимикробное вещество | Дозировка в диске, мкг | Диаметр зоны ингибирования роста, мм | ||
| Lactobacillus salivarius PC20b |
Lactobacillus curvatus PC723_5 |
Lactobacillus sp. PC18A | ||
| Пенициллин | 10 | 9,0 ± 0,2 | 7,0 ± 0,3 | 10,0 ± 0,3 |
| Эритромицин | 15 | 8,0 ± 0,3 | 7,0 ± 0,2 | 9,0 ± 0,3 |
| Линкомицин | 15 | 10,0 ± 0,3 | 10,0 ± 0,3 | 11,0 ± 0,3 |
| Цефалотин | 30 | 11,0 ± 0,3 | 9,0 ± 0,3 | 11,0 ± 0,2 |
| Тетрациклин | 30 | 10,0 ± 0,2 | 9,0 ± 0,2 | 11,0 ± 0,4 |
| Стрептомицин | 10 | 5,0 ± 0,2 | 4,0 ± 0,3 | 8,0 ± 0,3 |
| Ампициллин | 10 | 8,0 ± 0,3 | 5,0 ± 0,2 | 9,0 ± 0,2 |
Для повышения значения антибиотикоустойчивости исследуемых лактобацилл, их многократно культивировали на гидролизованном молоке с добавлением ранее используемых антибиотиков. В общей сложности было проведено 67 пассажей. Результаты антибиотикочувствительности микроорганизмов после пересевов представлены в таблице 33.
Таблица 33 – Антибиотикочувствительность молочнокислых микроорганизмов
| Антимикробное вещество | Дозировка в диске, мкг | Диаметр зоны ингибирования роста, мм | ||
| Lactobacillus salivarius PC20b |
Lactobacillus curvatus PC723_5 |
Lactobacillus sp. PC18A | ||
| Пенициллин | 10 | ‒ | ‒ | 3,0 ± 0,1 |
| Эритромицин | 15 | 2,0 ± 0,1 | 3,0 ± 0,1 | 4,0 ± 0,1 |
| Линкомицин | 15 | 5,0 ± 0,2 | 4,0 ± 0,1 | 5,0 ± 0,2 |
| Цефалотин | 30 | 4,0 ± 0,1 | 3,0 ± 0,1 | 4,0 ± 0,1 |
| Тетрациклин | 30 | 3,0 ± 0,1 | ‒ | 5,0 ± 0,1 |
| Стрептомицин | 10 | ‒ | ‒ | 3,0 ± 0,1 |
| Ампициллин | 10 | 2 ± 0,1 | ‒ | 4,0 ± 0,1 |
| Примечание: «‒» – отсутствие задержки роста. | ||||
В результате селекционного отбора удалось уменьшить зоны ингибиции роста исследуемых штаммов к используемым концентрациям антибиотиков как минимум в 2 раза, а в отдельных случаях они вообще не образовывали зон угнетения. Так, отсутствовала зона задержки роста отобранных культур при действии на Lactobacillus salivarius PC20b антибиотиков пенициллина и стрептомицина; Lactobacillus curvatus PC723_5 зона задержки роста отсутствовала при действии пенициллина, тетрациклина, стрептомицина и ампициллин.
Таким образом, за счет многократных пересевов отобранных культур удалось повысить их антибиотикоустойчивость, что обеспечит максимальный лечебно-профилактический эффект (пробиотический потенциал) разрабатываемого биопрепарата в промышленных условиях.
2.12 ИЗУЧЕНИЕ ПРОБИОТИЧЕСКОГО ПОТЕНЦИАЛА ИССЛЕДУЕМЫХ КУЛЬТУР ЛАКТОБАЦИЛЛ
Антагонизм к эталонной патогенной микрофлоре. Проводили изучение анагонистической способности изучаемых культур по отношению к эталонным тест-культурам патогенной и условно-патогенной микрофлоры методом отсроченного антагонизма (МУ 2.3.2.2789-10). Результы исследований представлены в таблице 34.
Таблица 34 − Антагонистическая активность изучаемых лактобактерий в отношении лабораторных тест-культур, мм
| Тест-культура | Изучаемые лактобактерии | ||
| Lactobacillus salivarius PC20b |
Lactobacillus curvatus PC723_5 | Lactobacillus sp. PC18A | |
| S. aureus АТСС 25923 | 20,0± 0,11 | 23,0± 0,21 | 22,0± 0,15 |
| В. subtilis 534 | 16,0± 0,09 | 13,0± 0,20 | 15,0± 0,16 |
| Е. coli АТСС 25922 | 23,0± 0,15 | 27,0± 0,19 | 22,0± 0,13 |
| Sh. sonnei 941 | 12,0± 0,14 | 8,0± 0,17 | 9,0± 0,17 |
| Р. aeruginosa АТСС 27853 | 21,0± 0,13 | 24,0± 0,20 | 18,0± 0,13 |
| Klebsiella pneumonia К1 5054 | 10,0± 0,16 | 26,0± 0,14 | 12,0± 0,15 |
| С. albicans АТСС 885-653 | 13,0± 0,13 | 15,0± 0,13 | 11,0± 0,16 |
В результате проведенных исследований установлено, что антагонистическая активность изучаемых культур лактобактерий в отношении эталонных тест-штаммов выше, чем при влиянии на полевые тест-культуры. Согласно рекомендациям, за положительный результат антагонистических свойств микроорганизмов принимается зона ингибирования ими тест-культуры в 20 мм (МУ 2.3.2.2789-10). В таком случае, высокие антагонистические свойства Lactobacillus salivarius PC20b проявляет в отношении S. aureus АТСС 25923 (20 мм), Е. coli АТСС 25922 (23 мм) и Р. aeruginosa АТСС 27853 (21 мм). Lactobacillus curvatus PC723_5 хорошо подавляет рост S. aureus АТСС 25923 (23 мм), Е. coli АТСС 25922 (27 мм), Р. aeruginosa АТСС 27853 (24 мм) и Klebsiella pneumonia К1 5054 (26 мм). К Lactobacillus sp. PC18A особо чувствительны S. aureus АТСС 25923 (22 мм) и Е. coli АТСС 25922 (22 мм). По отношению к другим эталонным тест-штаммам изучаемые лактобациллы также проявляют антагонистические свойства, но в меньшей степени, что не снижает их пробиотического потенциала.
Антагонизм к нормофлоре кишечника. Следующим этапом работы было выявить способность пробиотических штаммов ингибировать представителей нормофлоры кишечника, а также друг друга. Определение взаимодействия исследуемых штаммов с представителями нормофлоры изучали методом перпендикулярных штрихов. Проводили посев исследуемой культуры
в виде штриха по диаметру на чашку с питательной средой МРС. Перпендикулярно первому посеву делали штрихи других тест-культур. Культивирование проводили при 37 °С в течение 24−48 ч. Изучение способности штаммов-пробионтов in vitro ингибировать рост представителей нормофлоры кишечника, а также друг друга проводили, согласно рекомендациям (МУ 2.3.2.2789-10). В качестве тест-организмов применяли чистую культуру Lactobacillus acidophilus, выделенную из медицинского препарата «Линекс» (Словения); Lactobacillus acidophilus штамм n.v. Ep 317/402 (препарат «Наринэ», Армения); Lactobacillus galinarum ВКПМ-10134, выделенную из кишечника кур. Результаты исследований учитывали по зоне задержки роста (таблица 35).
Таблица 35 − Ингибирующая активность изучаемых лактобактерий
в отношении нормофлоры, мм
| Тест-культура | Зона ингибирования роста, мм | ||
| Lactobacillus salivarius PC20b |
Lactobacillus curvatus PC723_5 |
Lactobacillus sp. PC18A | |
| Lactobacillus acidophilus (шт. Линекс) |
0 | 0 | 0 |
| Lactobacillus acidophilus штамм n.v. Ep 317/402 |
0 | 0 | 0 |
| Lactobacillus galinarum ВКПМ-10134 |
0 | 0 | 0 |
| Lactobacillus salivarius PC20b | − | 0 | 0 |
| Lactobacillus curvatus PC723_5 | 0 | − | 0 |
| Lactobacillus sp. PC18A | 0 | 0 | − |
Результаты исследований показали, что при совместном выращивании культур в любом из вариантов, не было зафиксировано задержки роста исследуемых и тестируемых культур микроорганизмов.
Антиадгезивные свойства лактобацилл. Антиадгезивные свойства микроорганизмов – это способность культуры предотвращать или снижать прикрепительные действия (адгезию) патогенных микроорганизмов к объекту адгезии, в нашем случае эритроцитам крови барана. Изучаемое свойство лактобацилл определяли методом прямой и косвенной реакции (МУ 2.3.2.2789-10). Антиадгезивные свойства лактобактерий осушествляли по отношению
к тест-культурам (E. coli и S. aureus). Результаты исследований представлены в таблицах 36 и 37.
Таблица 36 – Результаты антиадгезивных свойств объектов исследований
к E. coli
| Метод исследований |
Объекты антиадгезивных свойств | Антиадгезивные свойства к контролю, % |
| Прямой метод |
Контроль | 100,0 |
| Lactobacillus sp. PC18A | 46,7 | |
| Lactobacillus salivarius PC20b | 41,5 | |
| Lactobacillus curvatus PC723_5 | 53,3 | |
| Косвенный метод |
Контроль | 100,0 |
| Супернатант Lactobacillus sp. PC18A | 34,3 | |
| Супернатант Lactobacillus salivarius PC20b | 37,2 | |
| Супернатант Lactobacillus curvatus PC723_5 | 29,8 |
В результате проведенных исследований установлено, что изучаемые культуры обладали по отношению к тест-штаммам антиадгезивными свойсвами, однако в различной степени. Так, при влиянии лактобацилл на адгезию E. coli к эритроцитам крови барана в прямом методе выявлено, что наибольшими антиадгезивными свойствами (53,3 %) обладала культура Lactobacillus curvatus PC723_5. Lactobacillus sp. PC18A и Lactobacillus salivarius PC20b способствовали антиадгезии E. coli равной 46,7 и 41,5 %, соответственно. При изучении антиадгезивных свойств супернатанта лактобактерий против E. coli, также выявлено положительное их действие. Супернатанты лактобактерий обладали антиадгезивными свойствами, что говорит о наличии метаболитов, подавляющих адгезию, которые выработали лактокультуры. Однако следует отметить, что использование живых культур лактобацилл обеспечивает более высокий результат антиадгезивных свойств.
Таблица 37 – Результаты антиадгезивных свойства объектов исследований
к S. aureus
| Метод исследований |
Объекты антиадгезивных свойств | Антиадгезивные свойства к контролю, % |
| Прямой метод |
Контроль | 100,0 |
| Lactobacillus sp. PC18A | 65,6 | |
| Lactobacillus salivarius PC20b | 54,1 | |
| Lactobacillus curvatus PC723_5 | 71,9 | |
| Косвенный метод |
Контроль | 100,0 |
| Супернатант Lactobacillus sp. PC18A | 36,8 | |
| Супернатант Lactobacillus salivarius PC20b | 42,3 | |
| Супернатант Lactobacillus curvatus PC723_5 | 31,4 |
Аналогичные результаты были получены при исследовании антиадгезивных свойств лактобактерий по отношению к S. aureus. Антиадгезивные свойства Lactobacillus sp. PC18A, Lactobacillus salivarius PC20b и Lactobacillus curvatus PC723_5 против S. aureus составили 71,9; 65,6 и 54,1 %, соответсвенно. Супернатанты исследуемых культур в отношении S. aureus также показали антиадгезивное действие, но в меньшей степени, чем нативные лактобациллы.
Адгезивные свойства лактобацилл (МУ 2.3.2.2789-10). Адгезивная активность – способность микроорганизмов прикрепляться к эпителиальным клеткам кишечника и размножиться прежде, чем энтероциты слизистого слоя будут обновлены. Данная способность микрофлорой обеспечивается за счет продукции ряда компонентов (пили или фимбрии), которые помогают закрепиться на клетке эпителия. Эти компоненты получили название – факторы колонизации или адгезии. Микроорганизмы могут экспрессировать различные типы пилий, которые кодируются разными хромосомальными и плазмидными генами. Это генетическое многообразие дает возможность культуре клеток приспосабливаться к меняющейся окружающей среде и использовать эту возможность по отношению
к различным поверхностным структурам организма-хозяина. Результаты адгезивности исследуемых лактобацилл по отношению к энтероцитам кишечника куриного эмбриона представлены в таблице 38.
Таблица 38 – Средний показатель адгезии (СПА) (при подсчете на 25
энтероцитах) исследуемых культур лактобактерий
| Исследуемые лактобациллы | Средний показатель адгезии (СПА) |
| Lactobacillus salivarius PC20b | 12,4 ± 0,51 |
| Lactobacillus curvatus PC723_5 | 17,5 ± 0,49 |
| Lactobacillus sp. PC18A | 14,1 ± 0,52 |
При подсчете количества микробных клеток, прикрепившихся к энтероцитам, установлено, что все исследуемые культуры лактобацилл обладали высокоадгезивными свойствами, так как показатель СПА варьировал от 12,4 до 17,5. При этом следует обратить внимание, что наибольшей адгезией к энтероцитам кишечника куриного эмбриона обладала культура Lactobacillus curvatus PC723_5 (СПА = 17,5), затем Lactobacillus sp. PC18A (СПА = 14,1) и Lactobacillus salivarius PC20b (СПА = 12,4).
Таким образом, результаты проведенных исследований показали, что выделенные из кишечника фазана молочнокислые культуры характеризуются высоким пробиотическим потенциалом, так как не обладают ингибирующим действием на нормофлору кишечника, в различной степени проявляют антагонистическую активность в отношении эталонной условно-патогенной тест-культуры, обладают высокими антиадгезивными и адгезивными свойствами,
в связи с чем, данные лактобациллы перспективны и могут быть включены
в состав биопрепарата микробного происхождения для применения в промышленном птицеводстве.
2.13 ИЗУЧЕНИЕ БЕЗОПАСНОСТИ ИССЛЕДУЕМЫХ ЛАКТОБАЦИЛЛ
Безопасность используемых в работе культур лактобактерий изучали путем проявления ими гемолитической активности на кровяном агаре и при пероральном введении их мышам (МУ 2.3.2.2789-10).
Определение гемолитической активности проводили на 5%-м кровяном агаре. Для его приготовления взвесь эритроцитов барана добавляли к охлажденной до 40−45 °С плотной питательной среде МРС с расчетом, чтобы взвесь эритроцитов составляла в агаре 5 %. Далее смесь осторожно перемешивали
и разливали в чашки Петри. После застывания среды проводили посев в разведениях культуры штамма для формирования отдельных колоний. Далее чашки инкубировали при 37 °С в течение 24 ч. Затем чашки Петри вынимали из термостата и осуществляли учет результатов. Положительным результатом гемолиза является формирование зоны лизиса.
Безвредность исследуемых культур in vivo изучали на лабораторных белых мышах с приблизительной массой 20 ± 1 г. Для этого колонии 24-часовой культуры лактобацилл, выращенных на агаризованной среде МРС, снимали
с агара петлей и суспендировали в 0,5 мл 0,9%-го раствора натрия хлорида до концентрации 100 млн/мл и выдерживали 30 мин при температуре 37 °С. По 0,5 мл полученной взвеси вводили через ротопищеводный зонд в желудок пяти белым мышам. Наблюдение за лабораторными животными проводили в течение пяти дней. Результаты исследований представлены в таблице 39.
Выработка гемолизинов считается фактором патогенности, в этой связи продукция гемолизина во многих случаях является маркером вирулентности.
В результате проведенного эксперимента, при выращивании лактокультур на 5 %-м кровяном агаре зоны лизиса вокруг них выявлено не было. Таким образом, результат гемолиза – отрицательный. Изучаемые культуры молочнокислых бактерий не проявляют гемолитической активности.
Таблица 39 – Безопасность исследуемых лактобацилл на мышах (n = 5)
| Группа | Вид животного |
Объем вводимой жидкости, способ введения | Результат испытаний, гол. | ||
| заболело | пало | выжило | |||
| Интактная | Мышь | − | 0 | 0 | 5 |
| Контрольная | 0,5 мл, перорально (физ. раствор) |
0 | 0 | 5 | |
| 1-я опытная | 0,5 мл, перорально
(Lactobacillus salivarius PC20b) |
0 | 0 | 5 | |
| 2-я опытная | 0,5 мл, перорально
(Lactobacillus curvatus PC723_5) |
0 | 0 | 5 | |
| 3-я опытная | 0,5 мл, перорально
(Lactobacillus sp. PC18A) |
0 | 0 | 5 | |
В результате проведенного эксперимента (в течении 5 дней) установлено, что во всех группах мышей не было зафиксировано гибели и заболевших животных. При ежедневном осмотре лабораторных животных можно было наблюдать, что мыши оставались подвижными и активными, регистрировалась удовлетворительная поедаемость корма, при сохранении всех жизненных рефлексов.
Таким образом, используемые в научно-иследовательской работе культуры лактобацилл, безопасны для применения.
2.14 ЭФФЕКТИВНОСТЬ ИСПОЛЬЗОВАНИЯ ПРОБИОТИКА ГАЛЛОБАКТ-Ф-ПЛЮС НА МОДЕЛИ ЭКСПЕРИМЕНТАЛЬНОГО ДИСБАКТЕРИОЗА У ЛАБОРАТОРНЫХ ЖИВОТНЫХ
Одним из главных предназначений клинического использования препаратов микробного происхождения является лечение и профилактика заболеваний, вызванных нарушением дисбаланса микробиоты желудочно-кишечного тракта. В связи с чем, следующим этапом исследований было изучение эффективности применения разработанного биопрепарата на модели экспериментального дисбактериоза кишечника у мышей и крыс, вызванного использованием антибиотика рифампицин. Для этого было изучено влияние пробиотика Галлобакт-Ф-Плюс и его отдельных компонентов (штаммов-пробионтов) в варианте лечебного и профилактического действия.
Оценку пробиотической эффективности использования консорциума выделенных культур в составе разработанного пробиотика Галлобакт-Ф-Плюс,
а также по отдельности проводили на модели экспериментального дисбактериоза у белых лабораторных беспородных мышей и крыс. Дисбаланс микробиоты вызывали введением перорально 0,5 см3 антибиотика рифампицин в дозировке 3,0 мг/гол. для мышей и 1,0 см3 рифампицина в дозировке 40,0 мг/гол. для крыс. Антибиотик задавали в течение 10 дней (Митрохин С. Д., Шендеров Б. А., 1989; Романчук Л. А., Елизбаращвили Д. А., Гинодман Г. А., 1991; Нуртдинова А. Н., 2003; Осипова И. Г., Васильева Е. А., 2006; Назырова Н. Р., 2007). Количество активных культур в дозировке препаратов составляло не менее 1,0 × 109 КОЕ/мл. Одновременно изучали лечебный и профилактичекий эффект от действия штаммов-пробионтов по отдельности и в составе пробиотика.
Для изучения лечебного эффекта используемых культур по отдельности
и в составе пробиотика Галлобакт-Ф-Плюс их задавали внутрижелудочно по 1,0 мл лабораторным мышам и крысам в течение 10 дней. Для этого было сформировано по каждому биообъекту исследования шесть групп-аналогов по 10 гол. в каждой: интактная группа – мыши и крысы получали стандартный рацион
и питье, без каких либо дополнительных препаратов и они были здоровы, ранее не получали антибиотик; контрольная группа – мыши и крысы с экспериментальным дисбактериозом, получали стандартный рацион, питье и физиологический раствор хлорида натрия; 1-я опытная группа – мыши и крысы с экспериментальным дисбактериозом, получали стандартный рацион и культуру Lactobacillus sp. PC18A; 2-я опытная группа – мыши и крысы с экспериментальным дисбактериозом, получали стандартный рацион и культуру Lactobacillus curvatus PC723_5; 3-я опытная группа – мыши и крысы с экспериментальным дисбактериозом, получали стандартный рацион и культуру Lactobacillus salivarius PC20b; 4-я опытная группа – мыши и крысы с экспериментальным дисбактериозом, получали стандартный рацион и пробиотик Галлобакт-Ф-Плюс.
Для изучения профилактического эффекта используемых культур по отдельности и в составе пробиотика Галлобакт-Ф-Плюс их задавали за 20 мин до введения рифампицина в течении 10 дней в аналогичных дозировках. Также были сформированы по каждому биообъекту исследования группы-аналоги по 10 гол. в каждой: интактная группа – здоровые мыши и крысы, получавшие стандартный рацион и питье, без каких либо дополнительных препаратов; контрольная группа – мыши и крысы получали в течении 10 сут рифампицин, а за 20 мин до введения антибиотика им задавали физиологический раствор хлорида натрия; 1-я опытная группа – мыши и крысы в течении 10 сут получали рифампицин, а за 20 мин до введения антибиотика им задавали Lactobacillus sp. PC18A; 2-я опытная группа – мыши и крысы в течении 10 сут получали рифампицин, а за 20 мин до введения антибиотика им задавали Lactobacillus curvatus PC723_5; 3-я опытная группа – мыши и крысы в течении 10 сут получали рифампицин, а за 20 мин до введения антибиотика им задавали культуру Lactobacillus salivarius PC20b; 4-я опытная группа – мыши и крысы в течении 10 сут получали рифампицин, а за 20 мин до введения антибиотика им задавали пробиотик Галлобакт-Ф-Плюс.
Изменение кишечного баланса микробиоты лабораторных животных оценивали по результатам микробиологических исследований фекалий толстой кишки и коэффициенту массы слепой кишки (КМСК) – отношение массы слепой кишки к массе тела лабораторного животного (Зельцер И. З., 1989; Назырова Н. Р., 2007).
Лечебное действие. Для выявления дисбиотического состояния микробного фона кишечника лабораторных животных после 10-дневнего введения антибиотика рифампицина проводили микробиологические исследования кишечника интактной и контрольной групп животных (таблица 40).
В результате введения антибиотика рифампицина у лабораторных животных контрольной группы были зафиксированы статистически достоверные признаки дисбактериоза кишечника. Выявлено, что в контрольной группе содержание полезных микроорганизмов (лакто- и бифидобактерий) снижалось по сравнению с условно-патогенной микрофлорой. Так, в контрольной группе мышей по сравнению с интактной группой титр лактобактерий с 7,75 снизился до 4,43 lg КОЕ/г; бифидобактрий – с 9,47 до 5,16 lg КОЕ/г (P < 0,05). При этом содержание эшерихий в контрольной группе увеличилось с 3,25 до 7,13 lg КОЕ/г, а условно-патогенных грибов – с 2,86 до 5,18 lg КОЕ/г при статистически достоверной разнице (P < 0,05). Также были выявлены изменения в показателе КМСК, значение которого в контрольной группе возрасло с 1,24 до 1,87 % (P < 0,05). Аналогичная картина выявлена в интактной и контрольной группах крыс.
Таблица 40 – Состояние кишечной микробиоты и коэффициент массы
слепой кишки (КМСК) у лабораторных животных интактной
и контрольной групп после 10-дневного применения
антибиотика
| Группа | Вид животного |
Содержание микробиоты кишечника, lg КОЕ/г | КМСК, % | |||
| лактобактерии | бифидобактерии | кишечная палочка | грибы | |||
| Интактная | Мышь | 7,75±0,11* | 9,47±0,09* | 3,25±0,10* | 2,86±0,07* | 1,24±0,01* |
| Крыса | 6,49±0,09* | 7,58±0,12* | 2,67±0,06* | 1,59±0,04* | 1,35±0,02* | |
| Контрольная | Мышь | 4,43±0,10 | 5,16±0,14 | 7,13±0,10 | 5,18±0,13 | 1,87 ±0,02 |
| Крыса | 3,12±0,07 | 3,65±0,08 | 6,78±0,09 | 5,22±0,09 | 2,03±0,02 | |
| Примечание:* – разница с контролем достоверна (P < 0,05). | ||||||
Таким образом, антибиотик рифампицин проявил мощное нарушение фекального микробиоценоза кишечника, тем самым спровоцировал дисбаланс
в пользу условно-патогенной микрофлоры, что проявилось признаком дисбактериоза.
Результаты влияния пробиотика Галлобакт-Ф-Плюс и его отдельных штаммов-пробионтов после 10-и дней терапии вызванного дисбактериоза, представлены в таблице 41.
В результате проведенных микробиологических исследований фекалий толстого отдела кишечника лабораторных животных была установлена статистически достоверная разницы между данными микробиоты контрольной группы и опытных. При этом статистически достоверные различия была зафиксированы во всех опытных группах и по всем изучаемым показателя в сравнении
с контрольной.
Таблица 41 – Состояние кишечной микробиоты и коэффициент массы
слепой кишки (КМСК) у лабораторных животных после
лечения экспериментального дисбактериоза
| Группа | Вид животного |
Содержание микробиоты кишечника, lg КОЕ/г | КМСК, % | |||
| лактобактерии | бифидобактерии | кишечная палочка | грибы | |||
| Интактная | Мышь | 7,75±0,11 | 9,47±0,09 | 3,25±0,10 | 2,86±0,07 | 1,24±0,01 |
| Крыса | 6,49±0,09 | 7,58±0,12 | 2,67±0,06 | 1,59±0,04 | 1,35±0,02 | |
| Контрольная | Мышь | 4,43±0,10 | 5,16±0,14 | 7,13±0,10 | 5,18±0,13 | 1,87 ±0,02 |
| Крыса | 3,12±0,07 | 3,65±0,08 | 6,78±0,09 | 5,22±0,09 | 2,03±0,02 | |
| 1-я опытная | Мышь | 6,54±0,10* | 7,67±0,07* | 4,34±0,05* | 3,67±0,05* | 1,46±0,02* |
| Крыса | 5,78±0,08* | 6,32±0,16* | 3,78±0,07* | 2,68±0,05* | 1,57±0,02* | |
| 2-я опытная | Мышь | 6,43±0,14* | 7,27±0,08* | 4,76±0,11* | 3,64±0,06* | 1,51±0,01* |
| Крыса | 5,23±0,10* | 5,87±0,11* | 3,71±0,07* | 2,48±0,06* | 1,58±0,01* | |
| 3-я опытная | Мышь | 5,98±0,06* | 7,21±0,16* | 4,55±0,10* | 3,48±0,04* | 1,49±0,02* |
| Крыса | 5,08±0,09* | 6,27±0,05* | 3,78±0,09* | 2,65±0,05* | 1,59±0,02* | |
| 4-я опытная | Мышь | 7,14±0,11* | 9,07±0,14* | 3,89±0,06* | 2,98±0,05* | 1,27±0,01* |
| Крыса | 6,14±0,13 | 7,15±0,08 | 2,97±0,06* | 1,85±0,03* | 1,39±0,02* | |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||||||
Применямые в научно-исследовательской работе лактобактерии по отдельности и в составе пробиотика Галлобакт-Ф-Плюс способствовали нормализации показателей кишечной микрофауны, а также КМСК у лабораторных животных. Наилучшая картина фекального микробиоценоза была выявлена при использовании в лечении лабораторных животных консоциума исследуемых лактобацилл в составе пробиотика Галлобакт-Ф-Плюс. Установлено, что применение пробиотика позволило повысить содержание лакто- и бифидобактерий, а также снизить уровень условно-патогенной микрофлоры у мышей, соответственно с 4,43 до 7,14 lg КОЕ/г; с 5,16 до 9,07 lg КОЕ/г; с 7,13 до 3,89 lg КОЕ/г и с 5,18 до 2,98 lg КОЕ/г (P < 0,05). Аналогично группам мышей наблюдалась статистически достоверная разница по отношению к контролю у опытных крыс – с 3,12 до 6,14 lg КОЕ/г; с 3,65 до 7,15 lg КОЕ/г; с 6,78 до 2,97 lg КОЕ/г и с 5,22 до 1,85 lg КОЕ/г (P < 0,05). Также следует обратить внимание, что опытная группа животных, получавших пробиотик Галлобакт-Ф-Плюс по всем изучаемым показателям (содержание лактобактерий, бифидобактерий, эшерийхий, грибов и КМСК) приближалась к значениям интактной группы (здоровые животные).
Таким образом, использование исследуемых культур лактобацилл, особенно в составе пробиотика Галлобакт-Ф-Плюс способствует высокому лечебному эффекту и дает основание предполагать, что применение данной композиции микроорганизмов позволит повысить эффективность лечения дисмикробных состояний и добиться стойких клинических эффектов.
Профилактическое действие. В связи с тем, что ведение промышленного животноводста, в том числе птицеводства, немыслимо без применения ряда антибиотиков, могут возникать случаи дисбактериальных нарушений, которые повышают риск заболеваний. В связи с этим, интересным было изучить возможность применения разработанного консоциума лактобактерий в составе пробиотика Галлобакт-Ф-Плюс для профилактики дисбиотический состояний при использовании антибиотиков. Для этого формировали из лабораторных мышей и крыс группы-аналоги согласно плану исследований.
Состояние микробиального фона кишечника у лабораторных животных после проведенной профилактики экспериментального дисбактериоза исследуемыми лактобактериями по отдельности и в составе пробиотика Галлобакт-Ф-Плюс представлены в таблице 42.
Таблица 42 – Состояние кишечной микробиоты и коэффициент массы
слепой кишки (КМСК) у лабораторных животных после
профилактики экспериментального дисбактериоза
| Группа | Вид животного |
Содержание микробиоты кишечника, lg КОЕ/г | КМСК, % | |||
| лактобактерии | бифидобактерии | кишечная палочка | грибы | |||
| Интактная | Мышь | 7,75±0,11 | 9,47±0,09 | 3,25±0,10 | 2,86±0,07 | 1,24±0,01 |
| Крыса | 6,49±0,09 | 7,58±0,12 | 2,67±0,06 | 1,59±0,04 | 1,35±0,02 | |
| Контрольная | Мышь | 4,67±0,11 | 5,11±0,12 | 7,06±0,09 | 5,23±0,10 | 1,92 ±0,01 |
| Крыса | 3,28±0,08 | 3,54±0,09 | 6,69±0,11 | 5,11±0,11 | 2,01±0,02 | |
| 1-я опытная | Мышь | 6,78±0,11* | 7,89±0,11* | 4,22±0,07* | 3,36±0,06* | 1,41±0,02* |
| Крыса | 5,83±0,09* | 6,64±0,11* | 3,54±0,09* | 2,35±0,04* | 1,52±0,01* | |
| 2-я опытная | Мышь | 6,58±0,11* | 7,48±0,17* | 4,48±0,10* | 3,42±0,08* | 1,51±0,01* |
| Крыса | 5,76±0,12* | 5,97±0,10* | 3,42±0,09* | 2,23±0,05* | 1,53±0,02* | |
| 3-я опытная | Мышь | 6,24±0,09* | 7,64±0,12* | 4,37±0,11* | 3,38±0,07* | 1,43±0,02* |
| Крыса | 5,32±0,08* | 6,38±0,19* | 3,41±0,07* | 2,45±0,06* | 1,51±0,01* | |
| 4-я опытная | Мышь | 7,43±0,15* | 9,22±0,12* | 3,47±0,07* | 2,95±0,06* | 1,25±0,02* |
| Крыса | 6,27±0,11 | 7,31±0,10 | 2,83±0,05* | 1,71±0,02* | 1,37±0,01* | |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||||||
Результаты изучения профилактического действия исследуемых культур по отдельности и в составе пробиотика Галлобакт-Ф-Плюс продемонстрировали положительную динамику в анализируемых показателях. При этом выявлено, что совместное использование антибиотика и физиологического раствора в контрольной группе дало отрицательный результат, так как в данной группе животных наблюдались дисбиотические нарушения в кишечнике, которые выражались в снижении полезной микрофлоры, повышении условно-патогенной, а также повышении показателя КМСК с 1,24 до 1,92 для мышей
и с 1,35 до 2,01 для крыс.
В пользу опытных групп по сравнению с контрольной наблюдались достоверно статистически значимые изменения в балансе кишечного микробиоценноза: повышался титр активных молочнокислых и бифидобактерий (P < 0,05), снижалась численность микроорганизмов группы кишечной палочки и грибов (P < 0,05), а также наблюдалось снижение показателя КМСК (P < 0,05). В целом, установлено, что применение в качестве профилактического средства композиции лактобактерий в составе пробиотика Галлобакт-Ф-Плюс показывает наилучшие результаты при анализе кишечного микробиома у лабораторных животных, которые близки к данным показателям
в группе здоровых животных.
Таким образом, результаты изучения лечебно-профилактического действия выделенных культур лактобацилл на модели экспериментального дисбактериоза кишечника у лабораторных животных показали, что применение их консоциума
в составе пробиотика Галлобакт-Ф-Плюс способствует стойкому клиническому результату, характеризующемуся повышением уровня представителей нормофлоры (лакто- и бифидобактерий), а также снижением численности представителей патогенной и условно-патогенной микробиоты, с последующим достижением уровня показателей характерных для здоровых животных.
2.15 ТОКСИКОЛОГИЧЕСКОЕ И РАЗДРАЖАЮЩЕЕ ДЕЙСТВИЕ ПРОБИОТИКА ГАЛЛОБАКТ-Ф-ПЛЮС
Токсикологическое действие пробиотика Галлобакт-Ф-Плюс оценивали путем изучения параметров острой, подострой и хронической токсичностей, а также раздражающих характеристик на подопытных лабораторных животных.
Изучение параметров токсичности пробиотика проводили согласно требований и рекомендаций (Тишков А. И., Аргунов М. Н., Ляшко Н. И., 1987; Хабриев Р. У., 2005; Смирнов А. А., Дорожин В. И., 2008).
Изучение кожно-резорбтивного действия и раздражающего свойства на слизистые оболочки глаз проводили на кроликах-альбиносах согласно ГОСТ Р ИСО 10993.10-99.
Изучение острой, подострой и хронической токсичностей проводили на клинически здоровых лабораторных животных (мыши, крысы), предварительно прошедших карантин в течение 14 дней. Лабораторные животные были распределены на группы по принципу пар-аналогов. Количество животных в каждой группе составляло по 10 гол. обоих полов, содержание самок и самцов было раздельным. В качестве лабораторных биообъектов исследований использовали нелинейных белых мышей с массой тела 18–20 г (возраст 2,5 мес) и половозрелых беспородных белых крыс с массой тела 180–200 г (возраст 2,5 мес). Лабораторные животные всех групп содержались в соотсветствии с требованиями (СП 2.2.1.3218-14).
При изучении показателей острой токсичности пробиотик лабораторным животным вводили однократно после 12-часовой голодной выдержки. Для получения достоверных результатов, эксперимент проводили в трех повторностях. Для перорального введения применяли металлический внутрижелудочный зонд. При введении препаратов животных фиксировали в вертикальном положении с незначительным запрокидыванием головы, вводили зонд в желудок
и медленно растворы. Кормление животных осуществляли через 3 ч с момента введения биопрепарата. После применения пробиотика за лабораторными животными вели наблюдение в течение двух недель, учитывая при этом изменения их общего поведения и состояния.
Титр микробных клеток в пробиотике Галлобакт-Ф-Плюс составлял 7,9 × 109 КОЕ/мл. С учетом массы лабораторных животных, а также рекомендаций по максимально вводимому объему жидкости зондовым способом (50,0 мл/кг) (ГОСТ Р ИСО 10993-11-2009) были рассчитаны объемы вводимые для мышей и крыс, которые составили – 1,0 и 10,0 мл пробиотика на одно животное, соответственно. Жидкости в объеме 10,0 мл крысам давали дробно
в течение суток.
Для изучения острой токсичности пробиотика было сформировано методом пар-аналогов пять групп животных: интактная группа – лабораторные мыши и крысы не получали дополнительных растворов, кроме стандартного поения и кормления; контрольная группа – вместо биопрепарата лабораторные мыши и крысы получали физиологический раствор в максимальном объеме (1,0 и 10,0 мл, соответственно) как в 3-й опытной группе; 1-я опытная группа – пробиотик в испытуемой дозе вводили перорально в объеме 0,2 мл мышам и 2,0 мл крысам; 2-я опытная группа − мышам жидкий пробиотик вводили внутрижелудочно в объеме 0,5 мл, а крысам– 5,0 мл; 3-я опытная группа – Галлобакт-Ф-Плюс вводили внутрижелудочно мышам в объеме 1,0 мл, а крысам – 10,0 мл.
Изучение подострой и хронической токсичностей кормовой добавки дает возможность выявить длительное действие их основных компонентов на функциональное состояние отдельных органов, тканей и систем, а также их способность к кумуляции. Подострая токсичность длилась 3 мес, а хроническая –
6 мес (Смирнов А. А., Дорожин В. И., 2008). При изучении подострой и хронической токсичностей опытным лабораторным животным пробиотик вводили ежедневно, внутрижелудочно в дозировках, составляющих 1/10, 1/20 и 1/50 от максимально введенной в остром эксперименте. После разбавления в полученные дозы прибавляли физиологический раствор до объема, обеспечивающего комфортное введение испытуемых добавок подопытным животным. За лабораторными животными велось ежедневное наблюдение, при этом контролировали их сохранность, поведение, аппетит и общее состояние. В начале и конце эксперимента проводилось индивидуальное взвешивание лабораторных животных. На финальном этапе эксперимента лабораторные животные подвергались умерщвлению путем эвтаназии с целью изучения морфо-биохимических показателей крови, а также проводился анализ состояния их органов и тканей, путем патолого-анатомического вскрытия.
На двух половозрелых кроликах-альбиносах разного пола (масса тела 2,2‒2,5 кг) для пробиотика проводили изучение кожно-резорбтивного действия. Кроликам за сутки до проведения исследований выстригали шерсть на симметричных участках обоих боков для аппликации. Один из выстриженных участков являлся контролем. Размер обрабатываемого участка кожи составлял от 5,0 до 8,0 % поверхности тела лабораторного кролика-альбиноса. На ватные тампоны наносили 1,0 мл биопрепарата и прикладывали его на выстриженный участок кожи. Фиксировали ватный тампон марлевой повязкой. Местно-раздражающее действие биопрепарата на кожу оценивали после 4-часовой выдержки,
а далее через 24; 48 и 72 ч. Учитывали наличие покраснения, повышение чувствительности, припухлости, воспаления, возможное проявление эритемы
и отека, а также другие общие реакции организма лабораторного животного на изучаемые биопрепараты. Проводили оценку индекса первичного раздражения.
Изучение раздражающего действия разрабатываемой кормовой добавки на конъюнктиву проводили на двух половозрелых кроликах-альбиносах обоего пола (масса 2,9‒3,1 кг). Перед исследованиями проводили осмотр состояния глаз каждого лабораторного кролика-альбиноса. Для проведения эксперимента подопытным кроликам в нижний конъюнктивальный мешок правого глаза вводили 0,1 мл изучаемого биопрепарата. После введения испытуемого биопрепарата нижнее и верхнее веки смыкали и выдерживали в течение 2–3 с. Контролем служил левый глаз. Осмотр глаза лабораторных животных после инсоляции биопрепарата проводили через 1; 24; 48 и 72 ч. Учитывали следующее: состояние конъюнктивы, роговицы, радужной оболочки, наличие отека, а также других выделений из правого глаза. Оценивали индекс первичного раздражения.
Изучение острой токсичности. Изучение острой токсичности биопрепарата Галлобакт-Ф-Плюс осуществляли на нелинейных белых мышах и половозрелых беспородных белых крысах. Результаты острой токсичности пробиотика представлены в таблице 43.
В результате проведенного эксперимента установлено, что в опытных группах не зависимо от того, какие в опыте применялись дозы пробиотика, не было зафиксировано гибели и заболевших животных, как и в группах интактной и контрольной. При ежедневном осмотре лабораторных животных можно было наблюдать, что мыши и крысы оставались подвижными и активными, регистрировалась удовлетворительная поедаемость корма, при сохранении всех жизненных рефлексов. В изучаемом диапазоне доз пробиотика смертность
в опытных группах не выявлена, что говорит об отсутствии полулетальной дозы. Таким образом, пробиотическая добавка Галлобакт-Ф-Плюс в исследуемых микробных концентрациях не вызывает выраженного токсикоза у лабораторных животных, в связи с чем ее можно согласно ГОСТ 12.1.007-76, отнести
к 4-му классу опасности (малотоксичные вещества).
Изучение подострой токсичности. Так как определить полулетальную дозу пробиотика в остром опыте не удалось, в качестве основной дозы при изучении подострой токсичности был взят максимальный объем вводимого биопрепарата для мышей и крыс (1,0 и 10,0 мл). Опытным лаборторным животным пробиотическую добавку задавали внутрь жедудка в концентрациях, составляющих 1/10, 1/20 и 1/50 от максимально вводимой в остром эксперименте. Таким образом, мыши получали пробиотик в объеме, соответственно, 0,1; 0,05 и 0,02 мл (7,9 × 108; 3,9 × 108 и 1,5 × 108 КОЕ/мл), а крысы 1,0; 0,5 и 0,2 мл (7,9 × 109; 3,9 × 109
и 1,5 × 109 КОЕ/мл). Помимо опытных групп формировались также группы, в которых не использовали аналогичные препараты: интактная группа – лабораторные мыши и крысы не получали дополнительных растворов, кроме стандартного поения и кормления; контрольная группа – вместо биопрепарата лабораторные мыши и крысы получали физиологический раствор в максимальном объеме (1,0
и 10,0 мл, соответственно). Введение растворов осуществляли ежедневно, через зонд, перорально. Результаты определения подострой токсичности биопрепарата Галлобакт-Ф-Плюс представлены в таблице 44.
Таблица 43 – Острая токсичность пробиотика Галлобакт-Ф-Плюс на лабораторных животных (n = 10)
| Группа | Вид животного |
Объем вводимой жидкости, способ введения |
Доза пробиотика, КОЕ/мл |
Результат испытаний, гол. | ||
| заболело | пало | выжило | ||||
| Интактная | Мыши | − | − | 0 | 0 | 10 |
| Крысы | − | − | 0 | 0 | 10 | |
| Контрольная | Мыши | 1,0 мл физ. раствора, перорально | − | 0 | 0 | 10 |
| Крысы | 10,0 мл физ. раствора, перорально | − | 0 | 0 | 10 | |
| 1-я опытная | Мыши | 0,2 мл Галлобакт-Ф-Плюс, перорально | 1,6 × 109 | 0 | 0 | 10 |
| Крысы | 2,0 мл Галлобакт-Ф-Плюс, перорально | 1,5 × 1010 | 0 | 0 | 10 | |
| 2-я опытная | Мыши | 0,5 мл Галлобакт-Ф-Плюс, перорально | 3,9 × 109 | 0 | 0 | 10 |
| Крысы | 5,0 мл Галлобакт-Ф-Плюс, перорально | 3,9 × 1010 | 0 | 0 | 10 | |
| 3-я опытная | Мыши | 1,0 мл Галлобакт-Ф-Плюс, перорально | 7,9 × 109 | 0 | 0 | 10 |
| Крысы | 10,0 мл Галлобакт-Ф-Плюс, перорально | 7,9 × 1010 | 0 | 0 | 10 | |
Таблица 44 – Подострая токсичность пробиотической добавки Галлобакт-Ф-Плюс на лабораторных животных (n = 10)
| Группа | Вид животного |
Объем вводимой жидкости, способ введения |
Доза пробиотика, КОЕ/мл |
Результат испытаний, гол. | ||
| заболело | пало | выжило | ||||
| Интактная | Мыши | − | − | 0 | 0 | 10 |
| Крысы | − | − | 0 | 0 | 10 | |
| Контрольная | Мыши | 1,0 мл физ. раствора, перорально | − | 0 | 0 | 10 |
| Крысы | 10,0 мл физ. раствора, перорально | − | 0 | 0 | 10 | |
| 1-я опытная | Мыши | 0,1 мл Галлобакт-Ф-Плюс, перорально | 7,9 × 108 | 0 | 0 | 10 |
| Крысы | 1,0 мл Галлобакт-Ф-Плюс, перорально | 7,9 × 109 | 0 | 0 | 10 | |
| 2-я опытная | Мыши | 0,05 мл Галлобакт-Ф-Плюс, перорально | 3,9 × 108 | 0 | 0 | 10 |
| Крысы | 0,5 мл Галлобакт-Ф-Плюс перорально | 3,9 × 109 | 0 | 0 | 10 | |
| 3-я опытная | Мыши | 0,02 мл Галлобакт-Ф-Плюс, перорально | 1,5 × 108 | 0 | 0 | 10 |
| Крысы | 0,2 мл Галлобакт-Ф-Плюс, перорально | 1,5 × 109 | 0 | 0 | 10 | |
Из эксперимента по наблюдению за общим состоянием и поведением животных установлено, что пробиотик в изучаемых дозировках не оказал негатичного влияния на организм подопытных биообъектов. Подопытные животные удовлетворительно переносили дозы биопрепарата. Гибели – не выявлено. Мыши и крысы в период постановки эксперимента оставались подвижными
и активными, а шерстный покров был гладким с характерным отблеском.
Проводилось изучение влияния пробиотика Галлобакт-Ф-Плюс на прирост живой массы лабораторных животных за период исследований. Учет живой массы мышей и крыс проводили перед постановкой опыта и после его завершения. Результаты эксперимента представлены в таблице 45.
Таблица 45 – Влияние Галлобакт-Ф-Плюс на рост и развитие лабораторных животных
| Группа | Масса тела, г | Прирост за опыт, г | |
| в начале опыта | в конце опыта | ||
| Мыши | |||
| Интактная | 18,87±0,27 | 25,63 ± 0,29 | 6,76 |
| Контрольная | 18,56±0,25 | 26,14 ± 0,25 | 7,58 |
| 1-я опытная | 18,37±0,28 | 31,89 ± 0,24* | 13,89 |
| 2-я опытная | 18,43±0,30 | 30,58 ± 0,27* | 12,15 |
| 3-я опытная | 18,69±0,27 | 31,43 ± 0,26* | 12,74 |
| Крысы | |||
| Интактная | 189,75±1,67 | 222,63 ± 1,85 | 32,88 |
| Контрольная | 190,12±1,58 | 224,36 ± 1,71 | 34,24 |
| 1-я опытная | 189,65±1,72 | 235,93 ± 1,68* | 46,28 |
| 2-я опытная | 189,71±1,59 | 234,71 ± 1,75* | 45,00 |
| 3-я опытная | 190,42±1,64 | 234,32 ± 1,63* | 43,90 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||
Установлено, что масса тела лабораторных животных, в частности 1–3-й опытных группах была выше, чем в контрольной группе при статистичеки достоверной разнице (Р < 0,05). Так, в 1-й опытной группе масса тела мышей была выше, чем в контрольной и интактной группах на 22,1 и 24,4 %, во 2-й опытной группе выше на 16,9 и 19,3 %, а в 3-й группе – на 20,2 и 22,6 %, соответсвенно.
Аналогичная тенденция наблюдалась в опытных группах лабораторных крыс. Так, в 1–3-й опытных группах живая масса крыс была достоверно выше, чем в контрольной на 5,2; 4,6 и 4,4 %, соответсвенно (Р < 0,05).
Проводилось изучение действия пробиотической добавки Галлобакт-Ф-Плюс на обмен веществ организма подопытных групп лабораторных мышей
и крыс через их морфо-биохимические показатели крови (таблица 46 и 47).
Таблица 46 – Морфологические и биохимические показатели крови
лабораторных мышей после применения пробиотика
Галлобакт-Ф-Плюс (n = 6)
| Показатель | Группа | ||||
| интактная | контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
|
| Морфологические показатели | |||||
| Лейкоциты, 109/л | 7,32±0,21 | 7,31±0,19 | 7,29±0,18 | 7,30±0,17 | 7,26±0,22 |
| Эритроциты, 1012/л | 8,21±0,17 | 8,32±0,16 | 8,53±0,12 | 8,51±0,19 | 8,49±0,20 |
| Тромбоциты, 109/л | 235,32±3,12 | 238,94±3,42 | 241,26±3,33 | 245,14±3,26 | 245,63±3,64 |
| Гемоглобин, г/л | 125,21±2,58 | 122,54±2,71 | 131,32±2,65 | 133,51±2,61 | 128,12±2,46 |
| Биохимические показатели | |||||
| Общий белок, г/л | 53,99±1,25 | 55,04±1,36 | 62,72±1,01* | 63,47±1,12* | 63,24±1,09* |
|
22,45±0,65 | 22,53±0,79 | 27,34±0,77* | 27,26±0,71* | 27,79±0,74* |
|
31,54±0,71 | 32,51±0,85 | 35,38±0,56 | 36,21±0,62 | 35,45±0,61 |
| А/Г коэффициент | 0,71±0,03 | 0,69±0,02 | 0,77±0,03 | 0,75±0,02 | 0,78±0,02 |
| Мочевина, мМ/л | 19,16±0,43 | 19,21±0,51 | 18,87±0,39 | 19,12±0,40 | 18,97±0,39 |
| Холестерин, мМ/л | 1,53±0,03 | 1,55±0,02 | 1,51±0,03 | 1,50±0,04 | 1,49±0,03 |
| Кальций, мМ/л | 2,37±0,05 | 2,31±0,04 | 2,38±0,04 | 2,41±0,05 | 2,35±0,03 |
| Фосфор, мМ/л | 1,32±0,03 | 1,36±0,02 | 1,42±0,03 | 1,49±0,04 | 1,43±0,02 |
| АСТ, Ед/л | 105,86±3,45 | 103,53±3,73 | 106,52±3,51 | 104,75±3,49 | 106,48±3,62 |
| АЛТ, Ед/л | 85,46±2,78 | 88,67±2,91 | 84,65±2,84 | 85,84±2,93 | 86,41±2,79 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
При анализе морфо-биохимического статуса крови лабораторных мышей после использования пробиотика Галлобакт-Ф-Плюс установлено, что в конце подострого эксперимента показатели крови находились в пределах физиологической нормы для данных биологических объектов исследований. Патологии
в обмене веществ выявлено не было. Однако по уровню отдельных показателей можно было выявить то, что использование добавки позитивно повлияло на организм лабораторных мышей. Так, в 1-й, 2-й и 3-й опытных группах мышей уровень эритроцитов и гемоглобина был более завышен, чем в контрольной группе на 2,5 и 7,1 %; 2,3 и 8,9 %; 2,0 и 4,6 %, соответственно. Повышение изучаемых показателей свидетельствует о том, что наблюдается улучшение насыщаемости органов и тканей подопытных животных, так как гемоглобин способен связыватся с кислородом, а эритроциты переносят его с током крови по организму, и как следствие наблюдается усиление окислительно-восстановительного процесса. Также повышеный в пределах нормы гемоглобин будет обеспечивать лучшее освобождение углекислого газа в легких. Содержание лейкоцитов в опытных группах было в пределах физиологической нормы, а также находилось на одном уровне с интактной и контрольной группой, что говорит об отсутствии негативных воздействий на организм лабораторных мышей от применяемой добавки, так как данные клетки крови ответственны за специфическую и неспецифическую защиту организма от внешних и внутренних негативных агентов. Аналогичная позитивная динамика была обнаружена при анализе биохимических показателей крови белых мышей. Статистически достоверная разница была выявлена при изучении уровня общего белка и его отдельных составляющих в крови опытных групп. Так, в 1-й опытной группе содержание общего белка было выше, чем в контроле, на 13,9 %, во 2-й – на 15,3 % и в
3-й ‒ на 14,9 % (Р < 0,05). Степень интенсивности процесса биосинтеза белковых компонентов крови показывает белковый коэффициент (А/Г), который в первой опытной группе был незначительно выше, чем в контрольной и интактной группах на 11,6 и 8,5 %, во 2-й опытной группе выше на 8,7 и 5,6 %, а в 3-й группе – на 13,0 и 9,8 %, соотвественно. По остальным биохимическим показателям крови существенной разницы не наблюдалось. В целом негативные последствия со стороны действия пробиотика Галлобакт-Ф-Плюс отсутствовали, а в отдельных случаях выявлено позитивное его влияние на обмен веществ в организме.
При проведении подострой токсичности на лабораторных крысах выявлено положительное влияние изучаемой добавки на морфо-биохимический статус крови животных. Из таблицы 47 видно, что содержание эритроцитов в цельной крови опытных групп животных было незначительно выше, чем в контрольной на 2,6; 1,9 и 1,7 %, соответственно. Уровень гемоглобина в эритроцитах по сравнению с контрольной группой в 1-й опытной группе был выше на 4,1 %, во 2-й – на 3,7 %, а в 3-й – на 6,3 %. Количество других элементов цельной крови находилось на одном физиологически нормальном уровне. В целом, морфологический статус крови лабораторных крыс показал отсутствие негативного воздействия пробиотика на организм животных, а даже наоборот, способствовал усилению обмена веществ.
Таблица 47 – Морфологические и биохимические показатели крови
лабораторных крыс после применения пробиотика
Галлобакт-Ф-Плюс (n = 6)
| Показатель | Группа | ||||
| интактная | контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
|
| Морфологические показатели | |||||
| Лейкоциты, 109/л | 11,34±0,35 | 11,41±0,29 | 11,31±0,28 | 11,33±0,33 | 11,29±0,31 |
| Гемоглобин, г/л | 102,63±3,78 | 105,73±3,62 | 110,11±3,71 | 109,63±3,84 | 112,42±3,67 |
| Тромбоциты, 109/л | 153,54±3,32 | 154,83±3,31 | 161,38±3,41 | 158,84±3,28 | 158,38±3,36 |
| Эритроциты, 1012/л | 5,32±0,12 | 5,37±0,13 | 5,51±0,11 | 5,47±0,15 | 5,46±0,17 |
| Биохимические показатели | |||||
| Общий белок, г/л | 66,13±1,34 | 65,13±1,21 | 68,80±1,36 | 68,51±1,31 | 69,60±1,27 |
|
29,64±0,82 | 28,98±0,78 | 31,12±0,76 | 31,02±0,79 | 31,78±0,81 |
|
36,49±0,74 | 36,15±0,69 | 37,68±0,54 | 37,49±0,59 | 37,82±0,61 |
| А/Г коэффициент | 0,81±0,02 | 0,80±0,01 | 0,83±0,02 | 0,83±0,02 | 0,84±0,03 |
| Мочевина, мМ/л | 5,64±0,16 | 5,60±0,19 | 5,58±0,15 | 5,62±0,13 | 5,61±0,14 |
| Холестерин, мМ/л | 1,87±0,03 | 1,83±0,04 | 1,79±0,03 | 1,81±0,03 | 1,81±0,03 |
| Кальций, мМ/л | 2,49±0,05 | 2,52±0,06 | 2,55±0,04 | 2,58±0,03 | 2,56±0,07 |
| Фосфор, мМ/л | 13,18±0,25 | 13,24±0,31 | 13,32±0,36 | 13,30±0,28 | 13,28±0,29 |
| АСТ, Ед/л | 153,65±3,85 | 155,48±3,67 | 158,54±3,74 | 156,38±3,48 | 156,21±3.54 |
| АЛТ, Ед/л | 135,17±3,45 | 139,24±3,21 | 139,49±3,67 | 141,82±3,42 | 138,15±3,33 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Аналогичная позитивная динамика была выявлена при анализе биохимических показателей крови подопытных крыс. Показатели крови располагались на уровне физиологической нормы. Показатель общего белка в сыворотке крови 1-й, 2-й и 3-й подопытных групп был выше, чем в группе контроля на 5,2
и 6,9 %. Негативного действия на биохимические показатели сыворотки крови лабораторных крыс не зафиксировано.
Таким образом, результаты подострой токсичности пробиотика Галлобакт-Ф-Плюс показали, что биопрепарат не оказал негативного (токсического) воздействия на организм подопытных лабораторных мышей и крыс.
Изучение хронической токсичности. Влияние пробиотика Галлобакт-Ф-Плюс на организм лабораторных животных в хроничеком эксперименте осуществляли аналогично подострому опыту, однако срок проведения испытаний составил 6 мес. Также дополнительно проводили патологоанатомическое вскрытие подопытных животных для изучения влияния биопрепарата на органы и ткани мышей и крыс.
Исследования показали, что лабораторные животные удовлетворительно переносили изучаемые дозы пробиотика. Гибели животных ‒ не выявлено. Нарушений в физиологической картине лабораторных животных не зафиксировано. Мыши и крысы в период проведения хронического опыта оставались жизнеспособны, движения были активные. Шерстный покров в норме (таблица 48).
Результаты изучения влияния действия биопрепарата на массу тела подопытных лабораторных животных, а также прирост представлены в таблице 49.
Из данных таблицы 49 установлено, что введение пробиотика Галлобакт-Ф-Плюс в рацион подопытных групп оказало статистически достоверное повышение на массу тела лабораторных животных. В первой, во второй и третьей опытных группах мышей масса животных на конец эксперимента была выше, чем в контрольной группе на 3,85 г (10,9 %), 3,56 г (10,1 %) и 3,25 г (9,2 %), соответственно (Р < 0,05). Прирост биообъектов был аналогично массе их тела, больше в опытных группах, чем в контрольной на 3,92 г (23,3 %), 3,54 г (21,0 %) и 3,3 г (19,6 %). В 1-й опытной группе лабораторных крыс наблюдаласть тенденция к возрастанию массы животных по отношению к крысам
в контрольной группе на 10,19 г, или 4,1 %. Во 2-й и 3-й опытных группах масса тела животных была достоверно выше, чем в контрольной группе на 11,75 г (4,8 %) и 10,3 г (4,2 %), соотвественно (Р < 0,05). Прирост в 1–3-й опытных группах крыс был выше, чем в группе контроля на 10,0 г, или 19,0 %.
Аналогично подострому эксперименту в хроническом опыте также проводили изучение влияния пробиотика Галлобакт-Ф-Плюс на организм лабораторных животных, через их морфологические и биохимические показатели цельной крови и сыворотки. Результаты исследований представлены в таблице 50 и 51.
Таблица 48 – Хроническая токсичность пробиотика Галлобакт-Ф-Плюс на лабораторных животных (n = 10)
| Группа | Вид животного |
Объем вводимой жидкости, способ введения | Доза пробиотика, КОЕ/мл |
Результат испытаний, гол. | ||
| заболело | пало | выжило | ||||
| Интактная | Мыши | − | − | 0 | 0 | 10 |
| Крысы | − | − | 0 | 0 | 10 | |
| Контрольная | Мыши | 1,0 мл физ. раствора, перорально | − | 0 | 0 | 10 |
| Крысы | 10,0 мл физ. раствора, перорально | − | 0 | 0 | 10 | |
| 1-я опытная | Мыши | 0,1 мл Галлобакт-Ф-Плюс, перорально | 7,9 × 108 | 0 | 0 | 10 |
| Крысы | 1,0 мл Галлобакт-Ф-Плюс, перорально | 7,9 × 109 | 0 | 0 | 10 | |
| 2-я опытная | Мыши | 0,05 мл Галлобакт-Ф-Плюс, перорально | 3,9 × 108 | 0 | 0 | 10 |
| Крысы | 0,5 мл Галлобакт-Ф-Плюс, перорально | 3,9 × 109 | 0 | 0 | 10 | |
| 3-я опытная | Мыши | 0,02 мл Галлобакт-Ф-Плюс, перорально | 1,5 × 108 | 0 | 0 | 10 |
| Крысы | 0,2 мл Галлобакт-Ф-Плюс, перорально | 1,5 × 109 | 0 | 0 | 10 | |
Таблица 49 – Влияние Галлобакт-Ф-Плюс на рост и развитие лабораторных животных
| Группа | Масса тела, г | Прирост за опыт, г | |
| в начале опыта | в конце опыта | ||
| Мыши | |||
| Интактная | 18,45±0,42 | 34,64 ± 0,47 | 16,19 |
| Контрольная | 18,54±0,44 | 35,37 ± 0,44 | 16,83 |
| 1-я опытная | 18,47±0,38 | 39,22 ± 0,42* | 20,75 |
| 2-я опытная | 18,56±0,39 | 38,93 ± 0,49* | 20,37 |
| 3-я опытная | 18,49±0,43 | 38,62 ± 0,44* | 20,13 |
| Крысы | |||
| Интактная | 195,32±2,87 | 245,56 ± 1,13 | 50,24 |
| Контрольная | 194,67±2,64 | 246,48 ± 1,11 | 51,81 |
| 1-я опытная | 195,14±2,83 | 256,67 ± 1,16 | 61,53 |
| 2-я опытная | 196,34±2,72 | 258,23 ± 1,22* | 61,89 |
| 3-я опытная | 194,89±2,79 | 256,78 ± 1,07* | 61,89 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||
Таблица 50 – Морфологические и биохимические показатели крови
лабораторных мышей после применения пробиотика
Галлобакт-Ф-Плюс (n = 6)
| Показатель | Группа | ||||
| интактная | контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
|
| Морфологические показатели | |||||
| Эритроциты, 1012/л | 8,03±0,12 | 8,10±0,14 | 8,34±0,14 | 8,39±0,13 | 8,38±0,12 |
| Гемоглобин, г/л | 102,76±2,87 | 105,38±2,94 | 110,34±2,89 | 108,36±2,77 | 109,83±2,92 |
| Тромбоциты, 109/л | 201,53±3,73 | 203,17±3,56 | 208,42±3,68 | 210,11±3,73 | 206,82±3,84 |
| Лейкоциты, 109/л | 7,21±0,22 | 7,20±0,18 | 7,19±0,19 | 7,22±0,16 | 7,22±0,18 |
| Биохимические показатели | |||||
| Общий белок, г/л | 51,25±0,76 | 52,73±0,67 | 59,62±0,81* | 60,57±0,80* | 59,65±0,75* |
|
21,13±0,79 | 21,26±0,75 | 25,14±0,81 | 25,75±0,79 | 24,96±0,83 |
|
30,12±0,71 | 31,47±0,69 | 34,48±0,68 | 34,82±0,71 | 34,69±0,75 |
| А/Г коэффициент | 0,70±0,02 | 0,67±0,03 | 0,72±0,03 | 0,74±0,02 | 0,72±0,03 |
| Холестерин, мМ/л | 1,67±0,03 | 1,62±0,04 | 1,53±0,03 | 1,55±0,04 | 1,54±0,03 |
| Мочевина, мМ/л | 21,46±0,52 | 20,89±0,49 | 21,64±0,45 | 20,84±0,46 | 20,17±0,43 |
| Кальций, мМ/л | 2,14±0,03 | 2,11±0,03 | 2,22±0,02 | 2,24±0,03 | 2,19±0,03 |
| Фосфор, мМ/л | 1,14±0,03 | 1,17±0,02 | 1,20±0,03 | 1,19±0,03 | 1,17±0,04 |
| АСТ, Ед/л | 125,27±3,56 | 122,02±3,73 | 126,28±3,84 | 124,27±3,63 | 123,87±3,81 |
| АЛТ, Ед/л | 91,63±1,76 | 90,46±1,85 | 91,47±1,68 | 90,97±1,73 | 89,68±1,95 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Таблица 51 – Морфологические и биохимические показатели крови
лабораторных крыс после применения пробиотика
Галлобакт-Ф-Плюс (n = 6)
| Показатель | Группа | ||||
| интактная | контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
|
| Морфологические показатели | |||||
| Эритроциты, 1012/л | 5,24±0,15 | 5,28±0,16 | 5,35±0,15 | 5,33±0,16 | 5,36±0,17 |
| Лейкоциты, 109/л | 11,01±0,19 | 11,24±0,19 | 10,57±0,18 | 11,03±0,18 | 10,96±0,21 |
| Тромбоциты, 109/л | 143,65±2,45 | 147,62±2,32 | 145,38±2,53 | 148,23±2,52 | 145,32±2,62 |
| Гемоглобин, г/л | 111,64±2,34 | 109,38±2,46 | 118,65±2,41 | 117,73±2,47 | 115,27±2,40 |
| Биохимические показатели | |||||
| Общий белок, г/л | 62,58±0,75 | 63,24±0,72 | 65,22±0,75* | 66,32±0,76* | 66,29±0,61* |
|
28,32±0,74 | 28,23±0,68 | 29,85±0,61 | 30,11±0,71 | 30,32±0,65 |
|
34,26±0,62 | 35,01±0,58 | 35,37±0,56 | 36,21±0,59 | 35,97±0,60 |
| А/Г коэффициент | 0,82±0,02 | 0,80±0,02 | 0,84±0,02 | 0,83±0,02 | 0,84±0,03 |
| Мочевина, мМ/л | 5,43±0,12 | 5,51±0,09 | 5,41±0,11 | 5,45±0,12 | 5,48±0,10 |
| Холестерин, мМ/л | 2,12±0,05 | 2,07±0,03 | 1,98±0,03 | 1,99±0,04 | 2,05±0,03 |
| Кальций, мМ/л | 2,23±0,04 | 2,27±0,03 | 2,31±0,03 | 2,28±0,04 | 2,27±0,03 |
| Фосфор, мМ/л | 11,26±0,22 | 11,35±0,25 | 11,38±0,19 | 11,33±0,23 | 11,29±0,22 |
| АСТ, Ед/л | 167,87±3,35 | 165,97±3,21 | 170,12±3,33 | 166,75±3,26 | 168,64±3,19 |
| АЛТ, Ед/л | 139,64±3,11 | 143,37±3,06 | 141,68±3,09 | 144,85±3,18 | 140,39±3,02 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Показатели крови всех изучаемых подопытных лабораторных животных были в норме для данных видов биообъектов. Патологии в изучаемых показателях, аналогично поставленным опытам в подостром эксперименте, не выявлено. Как в подостром, так и в хроническом опыте при использовании пробиотика Галлобакт-Ф-Плюс по анализируемым показателям крови можно было сделать вывод о позитивном влиянии добавки на организм мышей и крыс.
При проведении вскрытия лабораторных подопытных биообъектов, а также интактных и контрольных групп животных изменений и патологий в расположении и структуре внутренних органов – не выявлено. Органы в грудной
и брюшной полостях располагались анатомически правильно.
Таким образом, длительное применение пробиотической добавки Галлобакт-Ф-Плюс на нелинейных белых мышах и половозрелых беспородных белых крысах не оказало токсического воздействия на их организм.
Изучение кожно-резорбтивного и раздражающего свойства. Применение пробиотика предполагает его возможное попадание на кожу или слизистые оболочки глаз, в связи с чем, чтобы исключить какое-либо негативные вляиние в тахих случаях, важным было изучить раздражающие характеристики Галлобакт-Ф-Плюса на поверхность кожи и конъюнктиву глаз лабораторных животных. Биообъектами исследований выступали кролики-альбиносы.
Результаты накожных аппликаций показали, что в ходе опыта у лабораторных кроликов-альбиносов отсутствовали: местное покраснение, повышенная чувствительность, припухлость, воспаление, эритемы и отеки. Индекс первичного раздражения был зафиксирован как нулевой.
Конъюктивальная проба показала незначительное покраснение слизистой оболочки с одновременным выделением слезных желез сразу после введения биопрепарата, которые прошло в течении 2 мин. В дальнейшем негативного влияния не зафиксировано. Общее поведение и состояние животных через анализируемые промежутки времени (1; 24; 48 и 72 ч) оставалось положительным. Индекс первичного раздражения согласно ГОСТ Р ИСО 10993-10-2009 был равен нулю.
Таким образом, проведенный комплекс токсикологических исследований биопрепарата Галлобакт-Ф-Плюс доказал, что пробиотик не проявляет токсического и раздражающего свойства, в связи с чем может быть использован в промышленном птицеводстве независимо от дозы применения в изучаемых пределах.
2.16 ФАРМАКОЛОГИЧЕСКОЕ ДЕЙСТВИЕ ПРОБИОТИКА ГАЛЛОБАКТ-Ф-ПЛЮС НА ОРГАНИЗМ СЕЛЬСКОХОЗЯЙСТВЕННОЙ ПТИЦЫ
Изучение фармакологических свойств, а также выявление наиболее эффективной дозы и схемы применения пробиотика проводили на перепелах мясной породы Техасские белые и Фараон, а также цыплятах-бройлерах быстрорастущих кроссов Кобб 500 и Росс 308.
Для изучения влияния пробиотика Галлобакт-Ф-Плюс на организм перепелов был проведен научно-хозяйственный опыт. Схема эксперимента на перепелах представлена в таблице 52.
Таблица 52 – Схема научно-хозяйственного опыта на перепелах
при использовании пробиотика
| Группа | Кол-во голов | Условия кормления и выпаивания |
Схема выпаивания, дни |
| Контрольная | 200 | Стандартный рацион (СР) и питьевая вода (ПВ) |
|
| 1-я опытная | 200 | СР, ПВ + 0,25 мл/гол. Галлобакт-Ф-Плюс | 1; 3; 5; 7; 14; 21; 28; 35; 42; 49 |
| 2-я опытная | 200 | СР, ПВ + 0,5 мл/гол. Галлобакт-Ф-Плюс | |
| 3-я опытная | 200 | СР, ПВ + 0,75 мл/гол. Галлобакт-Ф-Плюс | |
| 4-я опытная | 200 | СР, ПВ + 1,0 мл/гол. Галлобакт-Ф-Плюс |
Для изучения влияния пробиотика Галлобакт-Ф-Плюс на организм цыплят-бройлеров быстрорастищих кроссов Кобб 500 и Росс 308 был проведен научно-хозяйственный опыт (таблица 53).
Продолжительность научно-хозяйственных экспериментов для каждой сельскохозяйственной птицы было индивидуальным. При выращивании перепелов испытания длились в течении 56 дней, для цыплят-бройлеров быстрорастущих кроссов – 42 дня.
Таблица 53 – Схема научно-хозяйственного опыта на цыплятах-бройлерах быстрорастищих кроссов при использовании пробиотика
Галлобакт-Ф-Плюс
| Группа | Кол-во голов | Условия кормления и выпаивания |
Схема выпаивания, дни |
| Контрольная | 60 | Стандартный рацион (СР) и питьевая вода (ПВ) |
|
| 1-я опытная | 60 | СР, ПВ + 0,25 мл/гол. Галлобакт-Ф-Плюс | 1; 3; 5; 7; 14; 21; 28; 35 |
| 2-я опытная | 60 | СР, ПВ + 0,5 мл/гол. Галлобакт-Ф-Плюс | |
| 3-я опытная | 60 | СР, ПВ + 0,75 мл/гол. Галлобакт-Ф-Плюс | |
| 4-я опытная | 60 | СР, ПВ + 1,0 мл/гол. Галлобакт-Ф-Плюс |
Каждый день велось наблюдение за клинико-физиологическим состоянием птицы путем осмотра поголовья, при этом обращали внимание на поведение, подвижность, перьевой покров, поедаемость корма и потребление воды. Во всех изучаемых группах рассчитывали процент сохранности птицы за весь период эксперимента. Еженедельно осуществляли изучение динамики живой массы птиц путем индивидуального взвешивания. По разнице начальной и конечной масс рассчитывали прирост птицы за весь эксперимент. Ежедневно велся учет за расходом комбикормов птицей. Конверсию корма рассчитывали как отношение съеденного комбикорма к 1 кг прироста за весь период исследований (Имангулов Ш. А., 2013).
Для изучения показателей мясной продуктивности после применения пробиотика в рационе птиц в конце исследований проводили их убой и анатомическую разделку. Проводили ветеринарно-санитарную экспертизу мяса птицы. Согласно требований изучали реакцию на пероксидазу, с формалином и сернокислой медью. Изучали содержание общего количества летучих жирных кислот (Правила ветеринарного осмотра, 1988). Проводили изучение показателя кислотности мяса птицы с использованием иономера pH-метр И-500 (ГОСТ
Р 51478-99). Органолептические показатели, упитанность, а также микробиологическую чистоту мяса птицы изучали согласно нормативной документации (ГОСТ Р 50396.1-92; ГОСТ Р 51944-2002; ГОСТ Р 54673-2011; ГОСТ 31962-2013). Дегустация мяса птицы и бульона из них проводилась согласно методическим рекомендациям (Имангулов Ш. А., 2013).
Для изучения переваримости компонентов комбикорма птицей проводился балансовый эксперимент за неделю до окончания исследований (Имангулов Ш. А., 2013). Химический анализ комбикорма и помета изучали по нормативной документации: отбор пробы – ГОСТ 26712-94; определение влаги – ГОСТ 13496.3-92; определение сухого остатка – ГОСТ 26713-85; определение сырого протеина – ГОСТ 13496.4-93; определение сырого жира – ГОСТ 13496.15-97; определение сырой золы – ГОСТ 26226-95; определение сырой клетчатки – ГОСТ 13496.2-91.
Изучался морфологический статус крови сельскохозяйственной птицы
и лабораторных животных. Количество клеток (эритроциты, лейкоциты и тромбоциты) в цельной крови подсчитывалось в камере с сеткой Горяева (Кудрявцев А. А., Кудрявцева Л. А., 1973). Содержание гемоглобина в эритроцитах измеряли на анализаторе НemoCue Hb. Биохимические показатели сыворотки крови подопытных животных и птиц изучали на полуавтоматическом настольном анализаторе Stat fax 4500. In vitro анализировали бактерицидную и лизоцимную активность сыворотки крови (БАСК и ЛАСК) согласно методикам (Смирнова Н. П., Кузьмина Т. А., 1966; Стогник В. И. [и др.], 1989).
Проводился анализ химического состава мышечной ткани птиц после использования биопрепаратов. Изучали содержание влаги по ГОСТ 9793-74; жира – ГОСТ 23042-78 и белка – ГОСТ 25011-81. Диетическое свойство мяса птиц определяли отношением количества жира к белку (индекс качества мяса) в соответствии с рекомендациями (Имангулов Ш. А., 2013). Аминокислотный скор мышц подопытных птиц изучался методом капиллярного электрофореза, на приборе «Капель–105 М», с применением предварительной гидролизацией белка мяса кислотным способом.
В эксперименте по эффективности использования пробиотика в рационе птиц после убоя проводили резекцию отделов кишечника для изучения микробного фона. Содержимое кишечника ресуспендировали в физиологическом растворе, а выделение и идентификацию микрофлоры осуществляли согласно методическим рекомендациям (Выделение и идентификация бактерий, 2004).
Расчеты экономической эффективности использования пробиотика на сельскохозяйственной птице осуществляли с учетом стоимости его расхода, количества съеденного комбикорма и сохранностью птицепоголовья. Учитывали фактическую стоимость комбикормов и цену продукции птицеводства в период проведения испытаний.
Полученные в опытах данные подвергались биометрической обработке
с помощью программного обеспечения фирмы Microsoft®. Критерий достоверности определяли по таблице Стъюдента. Разницу считали достоверной при Р < 0,05.
2.16.1 Фармакологическое действие пробиотика Галлобакт-Ф-Плюс на организм перепелов Техасской породы
Был проведен научный опыт на перепелах Техасской породы (мясной белый перепел), выведенной сотрудниками селекционно-генетического центра Всероссийского института птицеводства и ООО «Генофонд». Методом групп-аналогов в научно-исследовательской работе было сформировано пять групп птиц по 200 гол. в каждой: контрольная группа – птица получала стандартный рацион, рекомендованный ВНИТИП, и имела свободный доступ к воде; первая опытная группа – перепела получали стандартный рацион, а также дополнительно в воду вводился пробиотик Галлобакт-Ф-Плюс в дозе 0,25 мл/гол.; вторая опытная группа – аналогично первой, однако Галлобакт-Ф-Плюс вводили
в дозе 0,5 мл/гол.; третья опытная группа – объекты исследований получали стандартный рацион, рекомендованный ВНИТИП, а также в воду добавляли пробиотик в дозе 0,75 мл/гол.; четвертая опытная группа – объекты исследований получали стандартный рацион, а в воду добавляли пробиотик в дозе 1,0 мл/гол. Эксперимент длился в течение 56 дней.
Рост и развитие перепелов. В эксперименте использовались перепела
с признаками клинически здоровой птицы. Группы формировались методом аналогов. В течение всего эксперимента учитывали ряд зоотехнических показателей, результаты которых представлены в таблице 54 и рисунке 25.
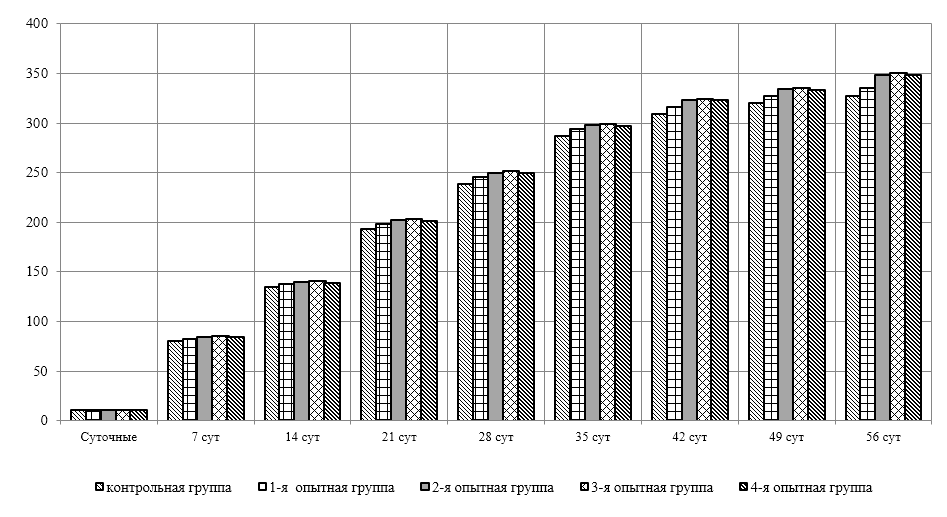
Рисунок 25 ‒ Динамика живой массы, г
Таблица 54 Зоотехнические показатели перепелов (n = 200)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Сохранность перепелов, % |
95,5 | 97,5 | 99,0 | 98,5 | 99,0 |
| Прирост живой массы за период выращивания (056 сут) | |||||
| Одной головы в среднем, г |
317,20 | 325,30 | 337,98 | 339,93 | 337,47 |
| Расход комбикорма за период выращивания (056 сут) | |||||
| 1-й головы, г | 1056,47 | 1087,95 | 1107,65 | 1121,12 | 1119,83 |
| Конверсия, кг | 3,33 | 3,34 | 3,28 | 3,29 | 3,31 |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
При изучении сохранности перепелов за период выращивания (56 дней) установлено, что наибольшая выживаемость птиц была зафиксирована в опытных группах. В частности, максимальный показатель был зафиксирован во второй и четвертой опытных группах, который составил 99,0 %, затем в третьей опытной группе – 98,5 % и далее в первой – 97,5 %. Самая низкая сохранность (95,5 %) наблюдалась в контрольной группе перепелов, при чем основная гибель перепелят была зафиксирована в первые недели жизни птицы.
При изучении живой массы перепелов установлено, что уже на первую неделю жизни при взвешивании была выявлена незначительная тенденция к повышению данного показателя в опытных группах. Так, в 1-й, 2-й, 3-й и 4-й опытных группах на 7-е сутки взвешивания показатель живой массы перепелов был незначительно выше, чем в контрольной на 2,6; 5,0; 5,6 и 4,6 %, соответственно. До 21-х суток наблюдалась аналогичная картина. Однако с 28-и дней
и до конца эксперимента во 2–4-й опытных группах была выявлена статистически достоверная разница по изучаемому показателю в сравнении с контрольной группой птиц. На 28-е сутки взвешивания перепелов установлено, что масса птиц во 2-й, 3-й и 4-й опытных группах была достоверно выше, чем в контрольной на 4,7; 5,5 и 4,8 % при Р < 0,05. При этом в 1-й опытной группе в данный период статистически достоверной разницы не выявлено, как и в принципе до конца эксперимента, но наблюдалась положительная динамика в изучаемом показателе в сравнении с контрольной группой. На 35-й день живая масса перепелов 2–4-й опытных групп была выше, чем в контрольной на 4,1; 4,5 и 3,8 % (Р < 0,05). Аналогичные статистически достоверные (Р < 0,05) показатели
в изучаемых опытных группах по отношению к контрольной были выявлены на 42-й и 49-й день взвешивания перепелов. На 56-е сутки были также зафиксированы достоверные различия во 2-й, 3-й и 4-й опытных группах
в сравнении с контрольной, соответственно, было выше на 6,3; 6,9 и 6,2 % (Р < 0,05). В 1-й опытной группе масса перепелов не значительно превышала контрольную на 2,5 %.
Расчет прироста перепелов во всех группах показал, что в опытных группах изучаемый показатель превосходил контрольную на следующие значения:
в 1-й опытной прирост выше на 2,6 %; во 2-й группе – на 6,6 %; в 3-й – на 7,1 % и в 4-й – на 6,4 %.
Одним из главных зоотехнических показателей эффективности использования в рационе животных, а в нашем случае птиц, добавок или биопрепаратов является показатель расхода кормов на 1 кг прироста массы (конверсия). Анализ данного значения показал, что в контрольной группе на 1 кг прироста требуется 3,33 кг комбикорма, в то время как в 1–4-й опытных группах необходимо 3,34; 3,28; 3,29 и 3,31 кг. Из этого следует, что наилучшее значение показателя было выявлено во 2-й опытной группе, которое было ниже, чем
в группе контроля, на 1,5 %. В 3-й опытной группе значение показателя было ниже, чем в контроле на 1,2 %, в 4-й опытной группе – на 0,6 %, а в 1-й опытной выше на 0,3 %.
Таким образом, перепела получавшие в своем рационе пробиотик Галлобакт-Ф-Плюс в изучаемых дозировках имели наилучшие зоотехнические показатели в сравнении с птицей без дополнительных биопрепаратов. Наилучшие показатели были зафиксированы в опытной группе, получавшей пробиотик Галлобакт-Ф-Плюс в дозе 0,5 мл/гол.
Морфологические и биохимические показатели крови. Такая система организма как кровь является одним из главных «посредников», отражающая большое количество процессов, которые происходят в тех или иных органах и тканях макроорганизма. Особая роль в крови отводится эритроцитам, тромбоцитам и лейкоцитам. Данные форменные элементы обладают большой функциональной способностью. Также следует обращать внимание на такой компонент крови как гемоглобин, который играет очень важную роль при переносе кислорода за счет эритроцитов от одного органа к другому, а также тканям. Наиболее глубокое понимание о влиянии на организм биопрепаратов дают результаты биохимических анализов сыворотки крови. Анализ данных компонентов крови позволяет выявить функциональное состояние внутренних органов, а также обнаружить развитие патологических и воспалительных явлений в организме.
В связи с этим, в середине (28-е сутки) и в конце (56-е сутки) опыта проводили забой птицы и взятие крови для изучения влияния пробиотика на организм птицы через ее морфологические и биохимические показатели. Данные анализа цельной крови и ее сыворотки представлены в таблицах 55 и 56.
Таблица 55 – Морфологический анализ крови перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| 28-е сутки | |||||
| Гемоглобин, г/л | 128,56±3,33 | 129,97±3,45 | 132,54±3,53 | 132,31±3,62 | 131,32±3,60 |
| Лейкоциты, 109/л | 23,65±0,64 | 23,12±0,54 | 22,43±0,52 | 22,65±0,29 | 22,15±0,49 |
| Тромбоциты, 109/л | 123,32±3,56 | 123,87±3,78 | 127,37±3,68 | 128,26±3,63 | 127,02±3,61 |
| Эритроциты, 1012/л | 3,65±0,10 | 3,71±0,10 | 3,75±0,10 | 3,76±0,11 | 3,74±0,11 |
| 56-е сутки | |||||
| Эритроциты, 1012/л | 3,42±0,08 | 3,47±0,10 | 3,52±0,11 | 3,51±0,11 | 3,50±0,08 |
| Тромбоциты, 109/л | 121,56±3,86 | 122,51±3,87 | 126,83±3,95 | 126,31±3,88 | 125,88±3,98 |
| Лейкоциты, 109/л | 26,34±0,54 | 25,58±0,42 | 24,73±0,49 | 24,67±0,48 | 24,71±0,64 |
| Гемоглобин, г/л | 127,54±3,94 | 128,87±3,87 | 131,76±4,01 | 130,87±3,99 | 130,79±3,87 |
Изучая морфологический состав отдельных форменных элементов крови, нами установлено положительное влияние использования в рационе перепелов пробиотика Галлобакт-Ф-Плюс, независимо от дозы и сроков применения. Так, на 28-й день количество эритроцитов в крови птиц 1–4-й опытных групп было незначительно выше, чем в контрольной на 1,6; 2,7; 3,0 и 2,5 %. Аналогичная динамика наблюдалась и на 56-й день изучения данного показателя, который был выше на 1,5; 2,9; 2,6 и 2,3 %, соответственно. Также незначительное положительное повышение наблюдалось тромбоцитов в опытных группах. Так, например, на 56-е сутки количество тромбоцитов по сравнению с контрольной группой было выше в 1-й опытной на 0,8 %; во 2-й – на 4,3 %; в 3-й – на 3,9 %
и в 4-й – на 3,6 %. Об отсутствии негативного влияния на организм птиц пробиотика также свидетельствует уровень лейкоцитов в крови птиц, содержание которого на 28-е и 56-е сутки анализа во всех группах было почти на одном уровне. Положительная динамика была зафиксирована в количестве гемоглобина в крови перепелов. Так, на 28-е сутки исследования цельной крови птиц
в 1–4-й опытных группах значение уровня гемоглобина было физиологически выше, чем в контрольной группе на 1,1; 3,1; 2,9 и 2,1 %, соответственно. На 56-й день исследований данного показателя была выявлена аналогичная картина.
В целом статистически достоверной разницы в разрезе изучаемых групп
не выявлено по анализируемым показателям крови. При этом введение в рацион опытных групп перепелов биопрепарата Галлобакт-Ф-Плюс способствовало незначительному физиологически нормальному повышению эритроцитов, тромбоцитов и гемоглобину, что обеспечило усиление обменных процессов
в организме птиц.
Результаты анализа сыворотки крови перепелов также продемонстрировали положительное влияние на организм птиц применяемого пробиотика (таблица 56). Проведенные исследования сыворотки крови перепелов в 28-дневном возрасте показали положительную динамику изучаемых показателей в опытных группах. Так, количество белка в крови 1–4-й опытных групп было выше, чем
в контрольной группе на 4,3; 10,6; 10,0 и 9,8 %. Количество белковых фракций в сыворотке крови перепелов опытных групп также было выше, чем в контрольной группе: альбуминов – на 5,3; 12,1; 11,1 и 11,4 %, а глобулинов – на 3,5; 9,5; 9,2 и 8,5 %, соответственно. Альбумино-глобулиновый коэффициент
в крови птиц 1-й, 2-й, 3-й и 4-й опытных групп превышал значение данного показателя в контрольной группе на 2,6; 3,9; 2,6 и 3,9 %. Аналогичные позитивные данные были получены при изучении других показателей сыворотки крови на 28-й день исследования без проявления статистически достоверной разницы. Однако на 56-й день эксперимента при анализе сыворотки крови перепелов была выявлена статистически достоверная разницы по отдельным изучаемым показателям. Так, во 2-й и 3-й опытных группах птиц в сыворотке крови белковый коэффициент (А/Г) был достоверно выше, чем в контрольной группе на 9,1
и 10,4 % (Р < 0,05).
Таблица 56 – Биохимический анализ сыворотки крови перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| 28-е сутки | |||||
| Общий белок, г/л | 33,23±0,87 | 34,65±0,91 | 36,76±1,02 | 36,56±0,99 | 36,48±1,01 |
| Альбумины, г/л | 14,56±0,32 | 15,33±0,40 | 16,32±0,42 | 16,17±0,38 | 16,22±0,45 |
| Глобулины, г/л | 18,67±0,54 | 19,32±0,48 | 20,44±0,51 | 20,39±0,53 | 20,26±0,58 |
| А/Г коэффициент | 0,77±0,01 | 0,79±0,02 | 0,80±0,01 | 0,79±0,02 | 0,80±0,01 |
| Холестерин, мМ/л | 3,52±0,08 | 3,48±0,09 | 3,38±0,09 | 3,40±0,07 | 3,40±0,10 |
| АСТ, ед./л | 354,24±7,54 | 352,18±8,02 | 347,67±7,86 | 347,54±8,13 | 348,38±7,69 |
| АЛТ, ед./л | 25,67±1,01 | 25,34±0,98 | 24,87±1,05 | 25,04±1,09 | 24,97±1,13 |
| Фосфор, мМ/л | 2,17±0,04 | 2,21±0,04 | 2,24±0,05 | 2,25±0,04 | 2,24±0,07 |
| Кальций, мМ/л | 1,85±0,05 | 1,94±0,03 | 2,00±0,04 | 2,01±0,03 | 1,98±0,04 |
| БАСК, % | 40,7±1,56 | 43,2±1,32 | 47,4±1,43 | 47,3±1,46 | 46,3±1,48 |
| ЛАСК, % | 31,2±1,05 | 33,4±1,15 | 35,1±1,09 | 35,6±1,08 | 35,2±1,10 |
| 56-е сутки | |||||
| Общий белок, г/л | 34,56±0,91 | 35,47±0,92 | 36,87±0,94 | 36,77±0,89 | 36,78±0,95 |
| Альбумины, г/л | 15,11±0,21 | 15,89±0,34 | 16,78±0,36 | 16,87±0,40 | 16,73±0,38 |
| Глобулины, г/л | 19,45±0,52 | 19,58±0,48 | 20,09±0,51 | 19,90±0,53 | 20,05±0,59 |
| А/Г коэффициент | 0,77±0,01 | 0,81±0,02 | 0,84±0,01* | 0,85±0,01* | 0,83±0,01 |
| Холестерин, мМ/л | 3,87±0,05 | 3,78±0,06 | 3,65±0,06* | 3,67±0,05* | 3,67±0,04* |
| АСТ, ед./л | 335,36±6,78 | 330,48±7,12 | 328,87±6,89 | 329,37±6,98 | 329,21±7,04 |
| АЛТ, ед./л | 24,61±1,02 | 24,12±0,97 | 23,96±1,01 | 24,08±0,94 | 24,04±0,97 |
| Фосфор, мМ/л | 2,19±0,01 | 2,25±0,02 | 2,29±0,01* | 2,29±0,01* | 2,28±0,01* |
| Кальций, мМ/л | 1,92±0,01 | 1,97±0,01 | 2,07±0,01* | 2,08±0,01* | 2,08±0,01* |
| БАСК, % | 43,1±0,86 | 47,4±0,92 | 52,3±1,02* | 53,1±0,98* | 52,8±0,96* |
| ЛАСК, % | 33,5±0,53 | 35,2±0,61 | 39,5±0,56* | 39,4±0,50* | 39,1±0,51* |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
Уровень холестерина в крови перепелов 2–4-й опытных групп был ниже, чем в контроле на 5,7; 5,2 и 5,1 % при статистически достоверном различии (Р < 0,05). Наблюдалась положительная динамика в изучении ферментов сыворотки крови перепелов опытных групп. Так, содержание АСТ в крови перепелов опытных групп было незначительно ниже, чем в контрольной группе на 1,5; 1,9 и 1,8 %. Уровень фермента АЛТ в сыворотке крови опытных групп было также незначительно ниже на 2,0; 2,6; 2,2 и 2,3 %. Была зафиксирована статистически достоверная положительная разница при изучении минерального обмена. Так, уровень фосфора и кальция в сыворотке крови птиц 2-й, 3-й и 4-й опытных групп был достоверно выше, чем в контрольной на 4,1 % (по фосфору), а также 7,8 и 8,3 % (по кальцию) (Р < 0,05). Уровень неспецифической резистентности организма птиц опытных групп был статистически достоверно выше в сравнении с показателями контрольной группы. Так, бактерицидная активность сыворотки крови 2-й, 3-й и 4-й опытных групп была выше, чем в контрольной на 9,2; 10,0 и 9,7 % (Р < 0,05). Лизоцимная активность во 2-й опытной группе была статистически выше, чем в контрольной, на 6,0 %, в 3-й опытной – на 5,9 %, а в 4-й – на 5,6 % (Р < 0,05).
Таким образом, применение в рационе птиц пробиотика Галлобакт-Ф-Плюс способствует повышению обменных процессов, в частности белкового
и минерального, при одновременном улучшении протекания жирового, за счет уменьшения холестерина в крови перепелов опытных групп, а также повышается иммунный статус перепелов. Негативное влияние от применения биопрепарата на морфобиохимический статус крови перепелов не выявлено.
Влияние пробиотика на пищеварение. Показатели переваримости компонентов корма в желудочно-кишечном тракте позволяют продемонстрировать эффективность использования биопрепаратов, в нашем случае пробиотика Галлобакт-Ф-Плюс. Анализ переваримости питательных компонентов комбикорма представлен на рисунке 26.
Проведенный балансовый эксперимент показал, что применение в рационе перепелов пробиотика Галлобакт-Ф-Плюс положительно сказывается на показателях переваримости компонентов комбикорма. Так, установлено, что в 1-й опытной группе переваримость органического вещества была выше, чем в контрольной группе на 2,9 %, во 2-й опытной – на 6,7 %, в 3-й группе – на 6,8 %,
а в 4-й – на 6,6 %. Количество сырого протеина птицей 1–4-й опытных групп усвоилось больше, чем в контрольной группе, на 2,9; 5,9; 6,0 и 5,7 %. Количество жира комбикорма перепелами опытных групп также переварилось больше, чем в контрольной группе на 3,1; 4,9; 5,1 и 4,7 %, соответственно. Сырая клетчатка, не смотря на то, что усваивается птицей незначительно, в опытных группах показатель ее переваримости был выше, чем в контрольной, на 3,1; 4,8; 5,0 и 4,8 %.
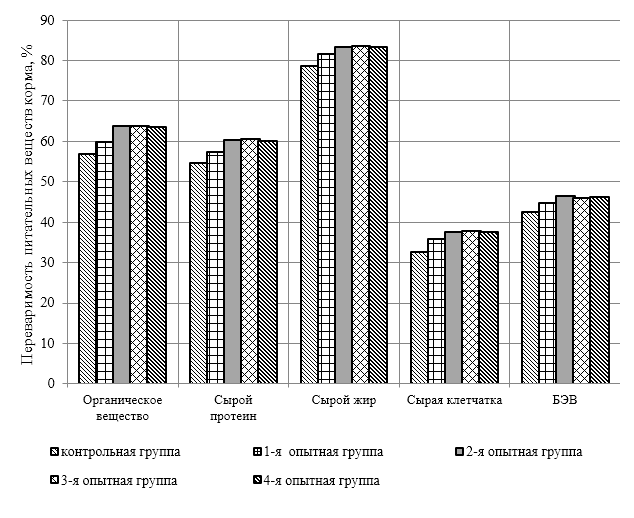
Рисунок 26 – Переваримость питательных веществ комбикорма, %
Также было выявлено незначительное повышение переваримости безазотистых экстрактивных веществ комбикорма перепелами 1–4-й опытных групп по сравнению с контрольной группой на 2,1; 3,9; 3,4 и 3,7 %, соответственно. Данные изменения носили динамический характер, статистически верной разницы не обнаружено.
Таким образом, пробиотик стимулирует процесс переваримости компонентов комбикорма, что способствует наилучшему росту живой массы экспериментальной птицы.
Микробиоценноз химуса кишечника. В случаях замедленного формирования микробного сообщества желудочно-кишечного тракта жизнеспособность сельскохозяйственной птицы может незначительно снижаться, что также может зависеть от санитарного состояния условий содержания и кормления. Данные изменения могут влиять на распространение таких желудочно-кишечных болезней как: гастроэнтерит, диспепсия, клоацит, энтероколит и токсико-септические инфекции. В связи с данными заболеваниями уместно в систему профилактических процедур включать применение биопрепаратов для формирования нормофлоры и колонизационной резистентности, к которым в первую очередь относят препараты микробного происхождения, в частности, пробиотики.
Для того чтобы изучить влияние пробиотика Галлобакт-Ф-Плюс на качественный и количественный состав микробной фауны ЖКТ перепелов, были проведены микробиологические анализы химуса слепых отростков кишечника исследуемой птицы. Результаты представлены в таблице 57.
Таблица 57 – Микробный баланс химуса кишечника перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Молочнокислые бактерии, 108 КОЕ/г |
1,2±0,03 | 2,8±0,03* | 4,2±0,03* | 4,8±0,04* | 4,5±0,03* |
| Бифидобактерии, 107 КОЕ/г | 0,9±0,03 | 2,1±0,04* | 2,7±0,04* | 2,8±0,04* | 2,9±0,03* |
| Эшерихии, 102 КОЕ/г | 4,5±0,05 | 3,2±0,03* | 2,5±0,04* | 2,3±0,03* | 2,2±0,04* |
| Энтерококки, 102 КОЕ/г | 1,1±0,02 | – | – | – | – |
| Псевдомонады, 10 КОЕ/г | – | – | – | – | – |
| Стафилококки, 102 КОЕ/г | 1,1±0,02 | 0,7±0,02* | 0,4±0,01* | 0,4±0,01* | 0,3±0,01* |
| Стрептококки, 102 КОЕ/г | 3,5±0,06 | 3,0±0,03* | 2,7±0,02* | 2,6±0,03* | 2,6±0,02* |
| Клостридии, 10 КОЕ/г | – | – | – | – | – |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Микробиологический анализ химуса слепых отростков перепелов после использования пробиотика Галлобакт-Ф-Плюс показал статистически достоверную разницу между опытными группами и контрольной. Так, при анализе содержания молочнокислых бактерий установлено, что в слепых отростках птиц опытных групп их титр был достоверно выше, чем в контрольной в 2,3; 3,5; 4,0 и 3,7 раза (P < 0,05). Количество активных бифидобактерий в 1–4-й опытных группах было выше, чем в контрольной, в 2,3; 3,0; 3,1 и 3,2 раза, соответственно, при статистически достоверной разнице (P < 0,05).
Было зафиксировано отсутствие в слепых отростках перепелов всех групп представителей клостридий и псевдомонад. Зафиксировано статистически достоверное снижение активных форм эшерихий в опытных группах перепелов
в сравнении с контрольной в 1,4; 1,8; 2,0 раза (P < 0,05). В опытных группах на питательных средах не было выявлено роста стрептококков, в то время как
в контрольной группе значение данного показателя наблюдалось. Также
в опытных группах было выявлено статистически достоверное снижения количеств представителей стафилококков и стрептококков.
Таким образом, пробиотик Галлобакт-Ф-Плюс способствует стимуляции роста полезной микрофлоры и снижению условно-патогенной микробиоты.
Мясная продуктивность, развитие внутренних органов и качество мяса перепелов. Результаты изучения мясной продуктивности перепелов после использования в их рационе пробиотика Галлобакт-Ф-Плюс в возрасте 56-и суток продемонстрированы в таблице 58.
В результате проведенного убоя установлено, что масса тушки после обескровливания во 2-й, 3-й и 4-й опытных группах оставалась статистически достоверно выше, чем в контрольной группе на 6,4; 6,7 и 6,2 % (Р < 0,05). Аналогичная тенденция наблюдалась и по другим показателям. Так, разница с контрольной группой была достоверна и при анализе непотрошеной тушки без перьев, масса которой во 2–4-й опытных группах превышала контроль на 6,4; 6,6 и 6,2 % (Р < 0,05). В 1-й опытной группе масса непотрошеной тушки без перьев была выше, чем в контрольной группе на 2,3 %, но разница не достоверна. Самые крупные мышцы тела у перепела – это грудные, масса которых была достоверно выше во 2-й, 3-й и 4-й опытных группах, чем в контрольной на 11,4; 11,6 и 10,6 % (Р < 0,05). Также во 2-й, 3-й и 4-й опытных группах была зафиксирована статистически достоверная разница по массе всех мышц тела птицы по отношению к контрольной группе, которая была выше на 13,1; 13,4 и 12,0 %, соответственно (Р < 0,05).
Таблица 58 – Мясная продуктивность перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Живая масса птицы перед убоем, г | 325,76±2,12 | 333,67±2,06 | 347,35±2,12* | 348,36±2,16* | 347,12±2,10* |
| Масса тушки после обескровливания, г | 316,31±2,01 | 324,32±2,02 | 336,58±2,06* | 337,56±2,11* | 335,87±2,02* |
| Масса непотрошеной тушки без перьев, г |
295,79±1,98 | 302,63±1,99 | 314,69±2,03* | 315,26±2,04* | 314,13±2,01* |
| Масса потрошеной тушки, г | 243,66±1,86 | 250,58±1,72 | 263,63±1,68* | 263,75±1,71* | 262,89±1,69* |
| Грудные мышцы, г | 70,68±0,71 | 73,45±0,63 | 78,76±0,75* | 78,87±0,74* | 78,14±0,70* |
| Масса бедренных мышц, г |
25,40±0,53 | 26,69±0,62 | 28,13±0,63 | 28,56±0,57 | 28,06±0,61 |
| Мышцы голени, г | 12,37±0,31 | 13,36±0,36 | 14,04±0,39 | 14,31±0,37 | 14,10±0,40 |
| Остальные мышцы, г |
12,05±0,45 | 12,67±0,42 | 14,71±0,47 | 14,87±0,48 | 14,56±0,43 |
| Всего съедобных мышц, г |
120,50±1,21 | 126,17±1,14 | 136,27±1,17* | 136,61±1,11* | 134,86±1,13* |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
Результаты влияния используемого биопрепарата на массу различных органов и накопление внутриполостного жира перепелов представлены в таблице 59.
Таблица 59 – Масса внутренних органов и внутриполосного жира, г (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Легкие | 3,32±0,10 | 3,35±0,11 | 3,38±0,09 | 3,38±0,10 | 3,37±0,08 |
| Печень | 8,35±0,33 | 8,38±0,29 | 8,39±0,30 | 8,40±0,31 | 8,39±0,27 |
| Сердце | 3,51±0,07 | 3,57±0,09 | 3,58±0,08 | 3,60±0,09 | 3,58±0,09 |
| Мышечный желудок | 6,17±0,21 | 6,26±0,19 | 6,34±0,19 | 6,37±0,18 | 6,35±0,17 |
| Железистый желудок | 1,18±0,03 | 1,22±0,04 | 1,26±0,04 | 1,28±0,04 | 1,25±0,03 |
| Кишечник | 11,29±0,34 | 11,37±0,33 | 11,43±0,32 | 11,44±0,34 | 11,41±0,34 |
| Внутриполостной жир | 2,57±0,07 | 2,47±0,09 | 2,21±0,08 | 2,22±0,09 | 2,25±0,07 |
Результаты взвешивания внутренних органов перепелов всех групп достоверных различий не показали. Разница массы отдельных органов была не значительна и не носила патологического характера, а была связана с изменением живой массы тела. В целом, масса внутренних органов была физиологически нормальной. При изучении массы внутриполостного жира статистически достоверной разницы не зафиксировано, однако следует отметить, что данный показатель в опытных группах имел тенденцию к снижению по сравнению с контрольной группой на 3,8; 14,0; 13,6 и 12,4 %, соответственно. По нашему мнению, часть жира птицей опытных групп, в связи с более активным метаболизмом, использовалось как источник энергии для поддержания обменных процессов.
Проводился анализ химического состава мышц перепелов всех групп
(таблица 60).
Таблица 60 – Химический состав мышц перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Ножные мышцы | |||||
| Влага, % | 72,18±1,24 | 71,84±1,08 | 71,74±1,18 | 71,72±1,26 | 71,70±1,28 |
| Белок, % | 22,43±0,51 | 23,11±0,51 | 23,35±0,46 | 23,36±0,42 | 23,30±0,53 |
| Жир, % | 4,87±0,14 | 4,53±0,11 | 4,38±0,12 | 4,39±0,10 | 4,38±0,14 |
| Зола, % | 0,52±0,02 | 0,52±0,01 | 0,53±0,02 | 0,53±0,01 | 0,53±0,02 |
| Индекс качества мяса (ИКМ) | 0,22 | 0,20 | 0,19 | 0,19 | 0,19 |
| Грудные мышцы | |||||
| Влага, % | 71,38±0,98 | 70,91±0,89 | 69,82±0,96 | 70,00±1,07 | 70,12±1,07 |
| Белок, % | 24,67±0,75 | 25,23±0,70 | 26,37±0,71 | 26,19±0,72 | 26,20±0,69 |
| Жир, % | 3,34±0,07 | 3,23±0,07 | 3,17±0,09 | 3,18±0,08 | 3,17±0,07 |
| Зола, % | 0,61±0,02 | 0,63±0,01 | 0,64±0,02 | 0,63±0,02 | 0,64±0,03 |
| Индекс качества мяса (ИКМ) | 0,14 | 0,13 | 0,12 | 0,12 | 0,12 |
Результаты изучения химического состава ножных и грудных мышц перепелов опытных групп продемонстрировали превосходство по сравнению с контрольной группой, но без достоверной разницы. Выявлено, что в мышцах перепелов опытных групп содержание влаги было незначительно ниже, чем в контрольной группе на 0,34; 0,44; 0,46 и 0,48 % (ножные), а также 0,47; 1,56; 1,38
и 1,26 % (грудные), соответственно. Количество белка в ножных мышцах перепелов 1–4-й опытных групп было выше, чем в контрольной на 0,68; 0,92; 0,93
и 0,87 %, а в грудных мышцах – 0,57; 1,7; 1,52 и 1,53 %, соответственно. Количество жира в мышцах птиц опытных групп незначительно снижалось по сравнению с контрольной. Важным показателем, характеризующим диетическое свойство мясной продукции, является индекс качества мяса, который
у перепелов в 1–4-й опытных группах был ниже, чем в контрольной группе на 9,1 и 13,6 % (ножные мышцы), а также 7,1 и 14,3 % (грудные мышцы),
соответственно.
Таким образом, использование пробиотика Галлобакт-Ф-Плюс способствует улучшению химического состав мышц птицы, при этом за счет снижения жира и повышения белка, а также снижения показателя индекса качества мяса повышается диетическое свойство мясной продукции и ее биополноценность.
Биополноценность мясной продукции перепеловодства также оценивалась содержанием особо значимых незаменимых аминокислот белка мышц птиц (рисунок 27).
Оценка аминокислотного скора мышц перепелов показала, что уровень изучаемых незаменимых аминокислот в опытных группах незначительно превосходил аналоги в контрольной группе. Установлено, что в ножных мышцах птиц 1-й опытной группы содержание лизина, триптофана, фенилаланина, лейцина и метионина было выше, чем в контрольной на 1,8; 3,8; 1,1; 1,1 и 1,7 %, соответственно. Во 2-й опытной группе данный показатель также был выше, чем в контрольной на 4,7; 7,6; 3,2; 2,0 и 2,3 %. Аналогичное превосходство зафиксировано в ножных мышцах 3-й и 4-й опытных группах в сравнении с контрольной и изучаемые аминокислоты были выше на 3,8; 7,3; 3,0; 1,6 и 2,6 %, а также 4,4; 7,6; 2,4; 1,4 и 1,7 % соответственно. В грудных мышцах птиц опытных групп содержание изучаемых незаменимых аминокислот также было выше, чем в контрольной группе. Так в 1−4-й опытных группах уровень лизина превосходил контрольную на 2,0; 4,2; 4,0 и 3,4 %, соответственно. Количество триптофана в 1-й опытной группе было выше, чем в контрольной группе на 2,9 %, во 2-й на 4,1 %, в 3-й опытной группе – 4,4 % и в 4-й – 3,2 %. В разрезе изучаемых групп в грудных мышцах была выявлена аналогичная картина по содержанию фенилаланина, лейцина и метионина.
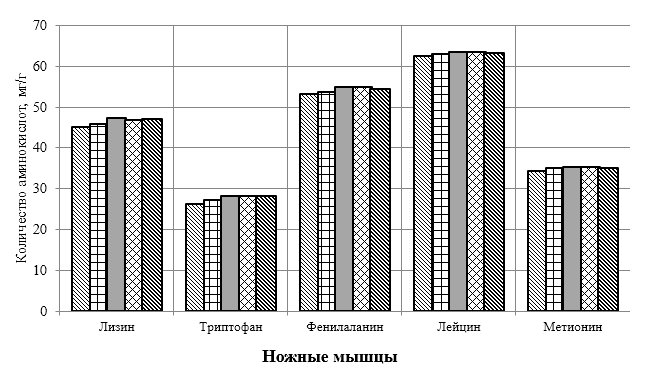
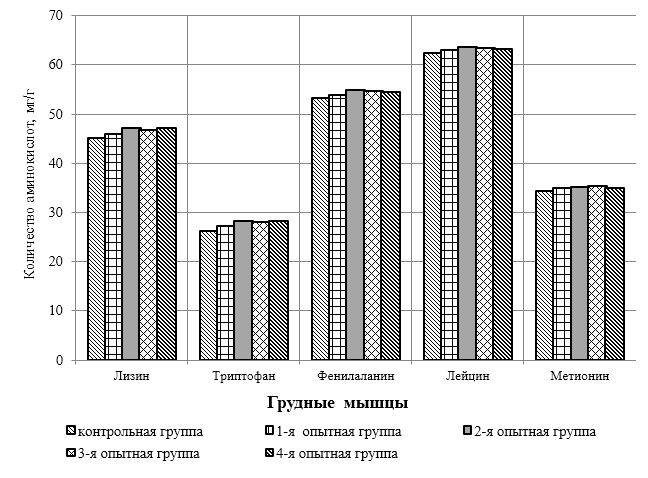
Рисунок 27 – Аминокислотный скор в мышцах перепелов, мг/г
Проводилась органолептическая и дегустационная оценка мяса перепелов и бульона из них. Органолептику изучали пробой варки. Для этого ножные
и грудные мышцы мелко измельчали, помещали в плоскодонную колбу, добавляли небольшое количество дистиллированной воды, закрывали колбу ватно-марлевой пробкой и кипятили на слабом огне в течение 30 мин. Как бульон закипал, с осторожностью приоткрывали пробку и определяли запах паров. Во всех пробах установлено, что запах оставался ароматным, свойственный мясу, иных ароматов не зафиксировано. Далее проводилась дегустация грудных, ножных мышц и бульона из них по пятибалльной шкале (таблица 61).
Таблица 61 − Дегустационная оценка мяса перепелов и бульона из них (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Ножные мышцы | |||||
| Аромат | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 |
| Вкус | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 | 4,8±0,1 |
| Нежность (жесткость) | 4,7±0,1 | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 |
| Сочность | 4,7±0,1 | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 |
| Грудные мышцы | |||||
| Аромат | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 |
| Вкус | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 | 4,8±0,1 |
| Нежность (жесткость) |
4,7±0,1 | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 |
| Сочность | 4,7±0,1 | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 |
| Бульон | |||||
| Аромат | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 |
| Вкус | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 | 4,8±0,2 |
| Прозрачность | 4,7±0,1 | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 |
| Крепость (наваристость) |
4,7±0,1 | 4,7±0,1 | 4,7±0,1 | 4,7±0,1 | 4,7±0,2 |
Результаты дегустационной оценки показали не существенную разницу
в разрезе изучаемых групп. Мясо перепелов всех групп имело коричнево-серый цвет, запах свойственный свежему мясу, приятный вкус. Бульон из-под мяса перепелов опытных и контрольных групп был прозрачный, с незначительным скоплением жира на поверхности. При этом дегустационной комиссией было отмечено, что мясо перепелов 1–4-й опытных групп было более сочным и нежным.
В целом, пробиотик Галлобакт-Ф-Плюс оказывает позитивное влияние на мясную продуктивность исследуемой птицы, а также способствует улучшению качества получаемой продукции перепеловодства.
Ветеринарно-санитарная экспертиза мяса птицы. Данные мероприятия являются обязательными при заключении о возможности дальнейшего использовании мяса животных, в том числе птицы, в рацион потребителя после применения того или иного биопрепарата.
Первоначально проводилось патологоанатомическое вскрытие птицы с целью выявления внешних признаков патологии. Вскрытие показало, что изменений в структуре органов и тканей перепелов не выявлено. У птиц всех групп органы имели правильное анатомическое расположение в полостях. Жидкость
в брюшной и плевральной полостях на зафиксирована. Просвет органов дыхания свободный, ткань легкого слабо-розового оттенка. Слизистая оболочка ЖКТ птиц серо-розового цвета без видимых повреждений и кровоизлияний.
В целом, патологические процессы не обнаружены.
Согласно ГОСТ Р 54673-2011 тушки перепелов испытуемых групп отнесены к 1-й категории. Так как по показателям упитанности тушки птиц имели хорошо развитую мускулатуру тела, грудь была округлой формы, наличие подкожного жир – умеренное. Масса переработанной тушки перепелов имела значение не ниже требований ГОСТа.
Обязательной процедурой при проведении ветеринарно-санитарной оценки является изучение физико-химических и микробиологических показателей мясо птицы (таблица 62).
Результаты исследований показали, что мясо перепелов всех групп было свежим и получено от здоровой птицы, так как бульон после добавления CuSO4 оставался прозрачным, при этом не было зафиксировано образования хлопьев или выпадения желеобразного сгустка сине-голубого или зеленоватого цвета.
При проведении реакции с формалином (формольная реакция) образование сгустка или хлопьев не выявлено, что говорило о свежести мяса перепелов. Следовательно, в мясе птиц всех групп не обнаружены продукты первичного метаболизма белка (аммиак и соли аммония).
Таблица 62 – Физико-химическая и микробиологическая оценка мяса
перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Р-ция с сернокислой медью | – | – | – | – | – |
| Р-ция с формалином | |||||
| Р-ция на пероксидазу | + | + | + | + | + |
| Кол-во ЛЖК, мг КОН/100 г | 2,15±0,07 | 2,12±0,05 | 2,10±0,06 | 2,10±0,07 | 2,12±0,08 |
| Количество микробных клеток в одном поле зрения микроскопа: |
|||||
| – с поверхности тушки | 3,35±0,11 | 3,30±0,10 | 3,29±0,09 | 3,30±0,10 | 3,28±0,11 |
| – с глубоких слоев | – | – | – | – | – |
| рН мяса, ед. | |||||
| Первые сутки | 6,81±0,21 | 6,72±0,21 | 6,70±0,18 | 6,68±0,17 | 6,75±0,15 |
| Вторые сутки | 6,52±0,20 | 6,41±0,26 | 6,26±0,20 | 6,20±0,21 | 6,29±0,22 |
| Третьи сутки | 6,05±0,17 | 5,96±0,25 | 5,84±0,16 | 5,83±0,24 | 5,91±0,18 |
При проведении реакции на наличие пероксидазы выявлено, что мясо было получено от здоровой птицы, так как вытяжка приобретала сине-зеленый цвет, переходящая в буро-коричневый в течение 1–2 мин.
При определении количества летучих жирных кислот в мясе перепелов выявлено, что во всех группах птиц количество ЛЖК находилось в пределах требований (до 4,5 мг КОН/100 г), что согласно утвержденной документации характерно для свежего и здорового мяса.
При изучении мазков-отпечатков с поверхности тушек перепелов под микроскопом во всех испытуемых группах птиц была обнаружена единичная микрофлора, преимущественно кокки. При микроскопии мазков-отпечатков глубоких слоев мышц птиц посторонняя микрофлора не зафиксирована, остатки распада ткани не выявлены, что подтверждает свежесть мяса перепелов.
При изучении pH выявлено, что такое резкое снижение показателя кислотности мышц в течение первых трех суток характерно для мяса полученного от здоровой птицы.
Таким образом, результаты проведенных исследований показали, что пробиотик Галлобакт-Ф-Плюс обладает стимулирующим действием на обмен веществ, повышает приросты, сохранность, способствует высоким показателям биополноценностии мясной продукции перепеловодства. Мясо перепелов может быть использовано в пищу независимо от дозы и сроков использования испытуемого биопрепарата.
В целом, результаты изучения фармакологических свойств пробиотика Галлобакт-Ф-Плюс, а также подбор эффективной его дозы показали, что по совокупности эффекта наилучшей дозой для перепелов можно считать 0,5 мл/гол. Данное количество пробиотика в рационе перепелов позволяет проявить высокий биоресурстный потенциал птицы.
Экономическая эффективность использования пробиотика. Важным показателем характеризующий эффективность применения тех или иных добавок для производства является экономическая целесообразность введения их в рацион животных, в частности, сельскохозяйственной птице. В связи с этим, для подбора наиболее экономически выгодной дозы с целью дальнейшей производственной апробации разработанных биопрепаратов, проводился предварительный расчет экономической эффективности их применения. Результаты экономической эффективности от использования пробиотика Галлобакт-Ф-Плюс на перепелах Техасской породы представлены в таблице 63.
В результате проведенной предварительной экономической оценки эффективности применения в рационе перепелов Техасской породы пробиотика Галлобакт-Ф-Плюс выявлено, что с учетом затрат на комбикорм и микробной добавки, прибыль в 1–4-й опытных группах составила 13 150,25; 14 241,30; 14 069,55 и 14 049,60 руб., против 12 562,30 руб. в контрольной группе, что выше на 587,95; 1 679,00; 1 507,25 и 1 487,30 руб., соответственно.
В целом, наилучшие результаты были получены во 2-й опытной группе,
в которой экономический эффект превышал контрольную группу на 13,36 %.
Таким образом, экономически целесообразно проводить производственную апробацию биопрепарата на перепелах Текасской породы в дозе, составляющей 0,5 мл/гол.
Таблица 63 – Экономическая эффективность использования пробиотической добавки Галлобакт-Ф-Плюс
| Показатель | Группа | |||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Поголовье в начале опыта, гол. | 200 | 200 | 200 | 200 | 200 | |
| Сохранность, % | 95,5 | 97,5 | 99,0 | 98,5 | 99,0 | |
| Поголовье в конце опыта, гол. | 191 | 195 | 198 | 197 | 198 | |
| Масса тушки на реализацию | ||||||
| Масса потрошенной тушки, г | 243,66±1,86 | 250,58±1,72 | 263,63±1,68* | 263,75±1,71* | 262,89±1,69* | |
| Масса потрошенной тушки всего, кг | 46,54 | 48,86 | 52,20 | 51,96 | 52,05 | |
| Затраты комбикорма за период выращивания | ||||||
| На голову, г | 1056,47 | 1087,95 | 1107,65 | 1121,12 | 1119,83 | |
| На все поголовье, кг | 201,79 | 212,15 | 219,31 | 220,86 | 221,72 | |
| Затраты пробиотика | ||||||
| Всего, мл | – | 487,5 | 990,0 | 1 477,5 | 1980,0 | |
| Экономическая эффективность | ||||||
| Цена 1 кг комбикорма, руб. | 30 | |||||
| Стоимость израсходованного корма всего, руб. | 6 053,70 | 6 364,50 | 6 579,30 | 6 625,80 | 6 651,60 | |
| Цена 1 л пробиотика, руб. | – | 60 | ||||
| Стоимость пробиотика всего, руб. | – | 29,25 | 59,40 | 88,65 | 118,80 | |
| Стоимость израсходованного корма и пробиотика, руб. | 6 053,70 | 6 393,75 | 6 638,70 | 6 714,45 | 6 770,40 | |
| Цена реализации 1 кг мяса перепелов, руб. | 400,0 | |||||
| Выручка от реализации мяса птицы, руб. | 18 616,00 | 19 544,00 | 20 880,00 | 20 784,00 | 20 820,00 | |
| Прибыль от реализации мяса птицы, руб. | 12 562,30 | 13 150,25 | 14 241,30 | 14 069,55 | 14 049,60 | |
| Экономический эффект от использования пробиотика к контролю,руб.% |
–
– |
587,95
4,68 |
1 679,00
13,36 |
1 507,25
12,00 |
1 487,30
11,83 |
|
2.16.2 Фармакологическое действие пробиотика Галлобакт-Ф-Плюс на организм перепелов породы Фараон
В качестве объектов исследований использовались перепела породы Фараон (французские), выведенные сотрудниками селекционно-генетического центра Всероссийского института птицеводства и ООО «Генофонд». Для проведения испытаний эффективности применения в рационе перепелов породы Фараон пробиотика Галлобакт-Ф-Плюс, а также подбора дозы биопрепарата, формировали группы птиц аналогично научным экспериментам на перепелах Техасской породы. Так, методом аналогов было сформировано пять групп птиц по 200 гол. в каждой: контрольная группа – птица получала стандартный рацион, рекомендованный ВНИТИП, и имела свободный доступ к воде; 1-я опытная группа – птица получала стандартный рацион, а также дополнительно в воду вводился пробиотик Галлобакт-Ф-Плюс в дозе 0,25 мл/гол.; 2-я опытная группа – пробиотик Галлобакт-Ф-Плюс вводили в дозе 0,5 мл/гол.; 3-я опытная группа – перепела получали стандартный рацион, а также в воду добавляли пробиотик
в дозе 0,75 мл/гол.; 4-я опытная группа – перепелам в воду вводили пробиотик Галлобакт-Ф-Плюс в дозе 1,0 мл/гол. Эксперимент длился в течение 56 дней.
Рост и развитие перепелов. Для проведения исследований использовалась здоровая птица. Результаты хозяйственных показателей перепелов после использования пробиотика представлены в таблице 64 и на рисунке 28.
Изучение сохранности птиц в процессе их выращивания (56 дней) показало, что наибольшая выживаемость птиц зафиксирована в экспериментальных группах. Так, наибольший уровень сохранности был зафиксирован во второй
и третьей опытных группах и составил 98,5 %, в первой и четвертой группах – 95,5 и 97,5 %, соответственно. Из данных таблицы 64 видно, что самый низкий показатель сохранности птицы (93,5 %) зафиксирован в контрольной группе.
Установлено, что показатели живой массы перепелов в течение первой недели жизни имели незначительную тенденцию к увеличению в опытных группах.
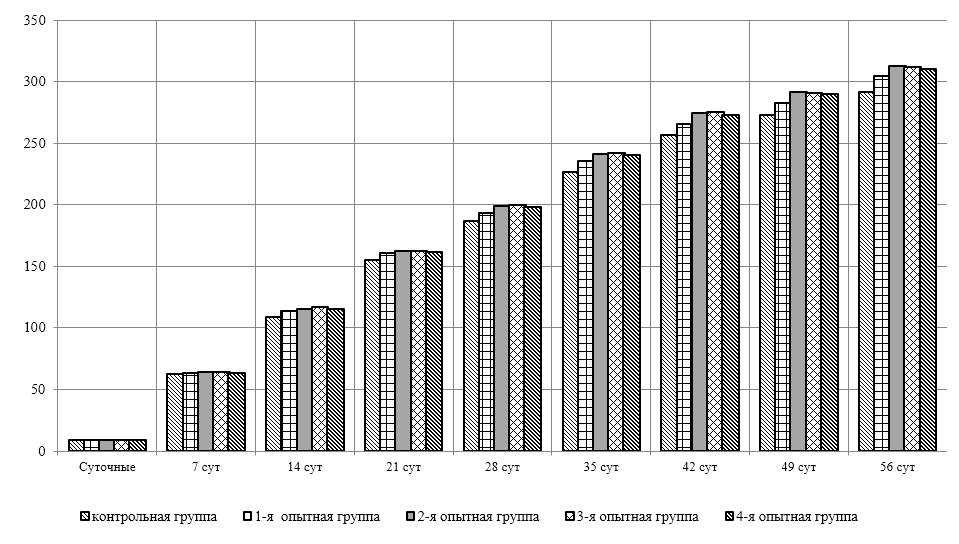
Рисунок 28 ‒ Динамика живой массы, г
Таблица 64 Хозяйственные показатели перепелов (n = 200)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Сохранность перепелов, % |
93,5 | 95,5 | 98,5 | 98,5 | 97,5 |
| Прирост живой массы за период выращивания (056 сут) | |||||
| Одной головы в среднем, г |
282,89 | 295,88 | 303,84 | 302,94 | 301,61 |
| Расход комбикорма за период выращивания (056 сут) | |||||
| 1-й головы, г | 937,85 | 980,37 | 990,40 | 993,65 | 989,49 |
| Конверсия, кг | 3,31 | 3,31 | 3,25 | 3,28 | 3,28 |
| * Разница с контролем достоверна (Р < 0,05) | |||||
Так, в 1-й, 2-й, 3-й и 4-й опытных группах на 7-е сутки выращивания данный показатель был незначительно выше, чем в контрольной на 1,2; 2,4; 2,9
и 1,7 %, соответственно. Аналогичная картина отмечалась до 35-х суток выращивания птицы. При этом к 42-дневному возрасту и до конца эксперимента
у перепелов 2–4-й опытных групп была установлена статистически достоверная разница по изучаемому показателю по отношению к контрольной группе птиц. При взвешивании перепелов на 42-е сутки установлено, что масса птиц во 2-й, 3-й и 4-й опытных группах была достоверно выше, чем в контрольной на 6,9; 7,4 и 6,5 % (Р < 0,05). При этом в 1-й опытной группе в данный период и до конца опыта статистически достоверной разницы не выявлено, но наблюдалась положительная динамика в изучаемом показателе в сравнении с контрольной группой. На 49-е сутки выращивания живая масса перепелов 2–4-й опытных групп была выше, чем в контрольной на 6,8; 6,6 и 6,4 % (Р < 0,05). На 56-й день были также зафиксированы достоверные различия во 2-й, 3-й и 4-й опытных группах в сравнении с контрольной и были выше на 7,2; 6,9 и 6,4 % (Р < 0,05), соответственно. В то время как масса перепелов 1-й опытной группы на 4,4 % превышала данные показатели контрольной.
При расчете прироста живой массы за весь период выращивания птицы установлено, что в опытных группах изучаемый показатель превосходит идентичный показатель в контрольной группе: в 1-й опытной группе – на 4,6 %; во 2-й группе – на 7,4 %; в 3-й – на 7,1 % и в 4-й – на 6,6 %.
Анализ конверсии показал, что в контрольной группе на 1 кг прироста требуется 3,31 кг комбикорма, в то время как в 1–4-й опытных группах необходимо 3,31; 3,25 и 3,28 кг. Из полученных данных следует, что оптимальное значение показателя было выявлено во 2-й опытной группе, что на 1,8 % ниже по отношению к показателям контроля. В 3-й и 4-й опытных группах значение показателя было ниже на 0,9 %. Показатели конверсии в контрольной и 1-й опытной группах идентичны.
Таким образом, перепела породы Фараон, в рационе которых применяли различные дозы пробиотика Галлобакт-Ф-Плюс, имели наилучшие хозяйственные показатели, чем у птиц контрольной группы. При этом максимальные значения изучаемых показателей были зафиксированы в группе птиц, где доза пробиотика составила 0,5 мл/гол.
Морфологические и биохимические показатели крови. Морфобиохимический статус птиц после применения исследуемого биопрепарата представлен
в таблицах 65 и 66.
Таблица 65 – Морфологический анализ крови перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| 28-е сутки | |||||
| Эритроциты, 1012/л | 3,72±0,08 | 3,72±0,06 | 3,74±0,05 | 3,73±0,08 | 3,74±0,10 |
| Тромбоциты, 109/л | 119,54±2,37 | 120,48±2,42 | 121,37±2,51 | 120,78±2,46 | 121,94±2,56 |
| Лейкоциты, 109/л | 21,63±0,45 | 21,36±0,64 | 21,44±0,54 | 21,62±0,58 | 21,86±0,46 |
| Гемоглобин, г/л | 124,83±2,48 | 124,76±2,61 | 126,48±2,52 | 125,62±2,42 | 126,13±2,57 |
| 56-е сутки | |||||
| Эритроциты, 1012/л | 3,68±0,05 | 3,78±0,04 | 3,86±0,06 | 3,84±0,05 | 3,83±0,07 |
| Тромбоциты, 109/л | 121,74±2,85 | 122,89±2,89 | 125,06±2,92 | 126,46±2,98 | 125,32±2,93 |
| Лейкоциты, 109/л | 22,78±0,78 | 22,36±0,65 | 21,58±0,57 | 21,74±0,48 | 22,16±0,67 |
| Гемоглобин, г/л | 125,68±2,53 | 126,87±2,61 | 128,84±2,49 | 128,37±2,57 | 127,89±2,72 |
При изучении влияния использования в рационе перепелов пробиотика Галлобакт-Ф-Плюс установлен положительный эффект на отдельные форменные элементы крови. Так, на 28-е сутки уровень эритроцитов в крови птиц 2-й, 3-й и 4-й опытных групп был незначительно выше, чем в контрольной на 0,5; 0,3 и 0,5 %, в то время как показатели контрольной и 1-й опытной группы находились на одном уровне.
Аналогичная динамика наблюдалась и на 56-й день изучения данного показателя во всех опытных группах, который был выше на 2,7; 4,9; 4,3 и 4,1 %, соответственно. Также наблюдалось незначительное положительное увеличение уровня тромбоцитов в опытных группах. Так, например, на 56-е сутки количество тромбоцитов по сравнению с контрольной группой было выше в 1-й опытной – на 0,9 %; во 2-й – на 2,7 %; в 3-й – на 3,9 % и в 4-й – на 2,9 %. Об отсутствии отрицательного влияния пробиотика на организм птиц также свидетельствует уровень лейкоцитов в крови, содержание которого на 28-е и 56-е сутки анализа во всех группах было почти на одном уровне. Также была установлена положительная динамика уровня гемоглобина в крови перепелов. Так, на 56-е сутки исследования цельной крови птиц в 1–4-й опытных группах значение уровня гемоглобина было физиологически выше, чем в контрольной группе на 0,9; 2,5; 2,1 и 1,8 %, соответственно.
Биохимические показатели крови занимают особое место и очень важны как для оценки физиологического статуса организма птиц, так и для своевременной диагностики патологических состояний. Результаты биохимического анализа сыворотки крови перепелов отражают то, что показатели крови птиц опытной группы проявили положительную динамику. Так, у перепелов, получавших корм с добавкой, уровень общего белка на 28-е сутки составил в 1-й опытной группе – 35,84 г/л; во 2-й – 36,32 г/л; в 3-й – 36,14 г/л; в 4-й – 35,95 г/л, что на 3,4; 4,8; 4,2 и 3,7 % выше, чем в контрольной группе. В возрасте 56-и дней по сравнению с предыдущим периодом проведения исследований содержание общего белка у перепелов повысилось во всех опытных группах с преобладанием концентрации показателя во второй группе по отношению к контролю на 9,3 %; в 1-й, 3-й и 4-й – на 3,5; 8,7 и 8,5 %, соответственно.
Из данных таблицы 66 видно, что количество альбуминов в сыворотке крови у перепелов на 28-е сутки во всех группах находилось приблизительно
в одном диапазоне – 15,63–15,87 г/л. Но уже к 56-м суткам выращивания наблюдалась тенденция к увеличению данного показателя в 1–4-й опытных группах по отношению к контролю на 4,8; 10,6; 9,9 и 9,3 %. Концентрация глобулинов
в крови перепелов на 28-е сутки в контрольной группе составил 19,04 ± 0,37 г/л, в 1-й опытной – 20,05 ± 0,41 г/л; во 2-й – 20,47 ± 0,39 г/л, в 3-й – 20,39 ± 0,43 г/л, в 4-й – 20,08 ± 0,48 г/л. В 56 дней наблюдалось увеличение количества глобулинов в контрольной и опытных группах. Так, у птиц контрольной и первой опытной группы показатель находился приблизительно на одном уровне, в то время как в крови перепелов 2–4-й групп наблюдалась тенденция к увеличению и было выше контроля на 6,6; 5,5 и 5,7 %. Таким образом, применение в рационе птицы пробиотика Галлобакт-Ф-Плюс, обуславливает увеличение уровня общего белка и его белковых фракций в опытных группах, что свидетельствует о положительном влиянии на уровень белкового обмена веществ, за счет чего будет происходить усиление естественной резистентности организма птицы.
Таблица 66 – Биохимический анализ сыворотки крови перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| 28-е сутки | |||||
| Общий белок, г/л | 34,67±0,96 | 35,84±0,83 | 36,32±0,74 | 36,14±0,69 | 35,95±0,84 |
| Альбумины, г/л | 15,63±0,46 | 15,79±0,49 | 15,85±0,52 | 15,75±0,39 | 15,87±0,42 |
| Глобулины, г/л | 19,04±0,37 | 20,05±0,41 | 20,47±0,39 | 20,39±0,43 | 20,08±0,48 |
| А/Г коэффициент | 0,82±0,02 | 0,79±0,01 | 0,77±0,01 | 0,77±0,02 | 0,79±0,02 |
| Холестерин, мМ/л | 3,42±0,07 | 3,37±0,08 | 3,32±0,08 | 3,31±0,09 | 3,32±0,07 |
| АСТ, ед./л | 343,54±5,67 | 342,43±6,09 | 342,53±5,81 | 341,74±6,11 | 342,54±5,65 |
| АЛТ, ед./л | 23,82±0,86 | 23,31±0,92 | 22,89±0,87 | 22,94±1,02 | 22,98±0,93 |
| Фосфор, мМ/л | 2,24±0,05 | 2,27±0,04 | 2,29±0,06 | 2,28±0,05 | 2,28±0,06 |
| Кальций, мМ/л | 1,75±0,03 | 1,80±0,04 | 1,83±0,03 | 1,84±0,04 | 1,83±0,03 |
| БАСК, % | 41,4±1,32 | 43,8±1,38 | 45,8±1,33 | 45,5±1,43 | 45,3±1,41 |
| ЛАСК, % | 32,3±0,87 | 33,7±0,97 | 34,2±0,94 | 34,1±0,96 | 33,8±1,02 |
| 56-е сутки | |||||
| Общий белок, г/л | 36,47±0,54 | 37,76±0,64 | 39,87±0,48 | 39,64±0,49 | 39,57±0,45 |
| Альбумины, г/л | 16,86±0,47 | 17,67±0,44 | 18,65±0,49 | 18,54±0,38 | 18,43±0,41 |
| Глобулины, г/л | 20,01±0,57 | 20,09±0,53 | 21,33±0,51 | 21,10±0,49 | 21,14±0,57 |
| А/Г коэффициент | 0,84±0,02 | 0,87±0,01 | 0,87±0,01 | 0,87±0,01 | 0,87±0,02 |
| Холестерин, мМ/л | 3,73±0,03 | 3,62±0,02 | 3,44±0,05* | 3,42±0,06* | 3,47±0,04* |
| АСТ, ед./л | 345,65±4,62 | 344,68±4,86 | 343,76±5,32 | 343,71±4,84 | 343,84±5,23 |
| АЛТ, ед./л | 24,27±0,76 | 24,03±0,92 | 23,86±0,96 | 24,02±0,91 | 23,93±0,89 |
| Фосфор, мМ/л | 2,27±0,01 | 2,35±0,02 | 2,43±0,01* | 2,45±0,01* | 2,41±0,01* |
| Кальций, мМ/л | 1,76±0,01 | 1,87±0,02 | 2,05±0,01* | 2,03±0,01* | 2,01±0,01* |
| БАСК, % | 42,4±0,46 | 45,3±0,53 | 52,8±0,63* | 52,5±0,58* | 51,8±0,56* |
| ЛАСК, % | 34,5±0,32 | 35,3±0,42 | 38,5±0,41* | 39,1±0,40* | 38,4±0,31* |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
К 56-м суткам у перепелов, потреблявших комбикорм с добавление Галлобакт-Ф-Плюс, отмечается статистически достоверное повышение уровня фосфора, кальций, БАСК, ЛАСК и снижение уровня холестерина. Так, уровень холестерина в крови перепелов опытных групп был ниже, чем в контроле: в 1-й – на 2,9 %; во 2-й – на 7,8 %; в 3-й – на 8,3 % и в 4-й – на 6,9 % при статистически достоверном различии (Р < 0,05). Это свидетельствует, что все процессы метаболизма липидов находятся в физиологической норме. Данному виду птицы свойственна высокая энергия роста, интенсивный обмен веществ, хорошо развитая воспроизводительная функция. Эти биологические особенности оказывают влияние на процессы минерального обмена. При изучении влияния биопрепарата Галлобакт-Ф-Плюс на организм перепелов из минеральных элементов в крови определяли кальций и фосфор. К 56-м суткам выращивания установлено статистически достоверное увеличение уровня фосфора и кальция во 2–4-й опытных группах по отношению к контролю: по фосфору – на 7,1; 7,9
и 6,2 %, по кальцию – на 16,5; 15,3 и 14,2 %. Полученные данные свидетельствуют об эффективном усвоении минеральных веществ птицей и активизации минерального обмена.
Влияние добавки на естественную резистентность организма оценивали по уровню бактерицидной и лизоцимной активности сыворотки крови (БАСК
и ЛАСК). Так, бактерицидная активность у птиц контрольной группы составила 42,4 %, а у перепелов, потреблявших корма с биопрепаратом, она достоверно
(Р < 0,05) превышала во 2-й группе на 10,4; в 3-й – на 10,1 и в 4-й – на 9,4 %. Показатель лизоцимной активности сыворотки крови статистически был выше во 2–4-й опытных группах, чем в контрольной на 4,0; 4,6 и 3,9 %. Следовательно, уровень БАСК и ЛАСК у птиц опытных групп свидетельствует о повышенной способности организма к самоочищению.
Таким образом, исследуемый пробиотик не оказывает негативного влияния на организм перепелов, стимулирует различные виды обмена веществ,
в частности белковый – за счет повышения общего белка и белковых фракций, жировой – за счет снижения холестерина в крови, минеральный – за счет незначительного повышения кальция и фосфора в сыворотки крови перепелов, а также наблюдается повышение иммунного статуса птицы – за счет повышения показателей бактерицидной и лизоцимной активностей.
Влияние пробиотика на пищеварение. Показатели переваримости питательных компонентов комбикорма после применения пробиотика представлены на рисунке 29.
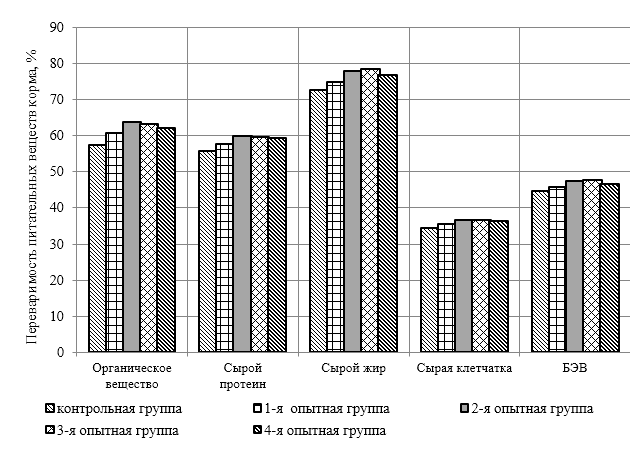
Рисунок 29 – Переваримость питательных веществ комбикорма, %
В ходе эксперимента было выявлено, что введение в рацион Галлобакт-Ф-Плюс положительно сказалось на переваримости питательных веществ комбикорма птицей. Установлено, что усвояемость органического вещества комбикорма в контрольной группе составило 57,43 %, в то время как в опытных группах наблюдалась тенденция к увеличению данного показателя: в 1-й группе – на 3,2 %; во 2-й – на 6,4 %; в 3-й – на 5,9 % и в 4-й – на 4,8 %. Важным показателем корма является протеиновая питательность, которая удовлетворяет потребность птицы во всех заменимых и незаменимых аминокислотах. Из данных рисунка 29 установлено, что процент переваримости сырого протеина корма лучше в 1–4-й опытных группах и превышает контроль на 1,8; 4,1; 3,9
и 3,5 %. Птица экспериментальных групп также лучше переваривала сырую клетчатку потребляемого корма, чем перепела контрольной группы, в среднем на 1,8 %.
Переваримость сырого жира комбикорма в контрольной группе составило 72,78 % против 74,82 % в 1-й опытной группе, 77,98 – во 2-й; 78,43 – в 3-й и 76,89 % – в 4-й, что выше на 2,0; 5,2; 5,7 и 4,1 %, соответственно. Что касается переваримости БЭВ, то у перепелов 1–4-й сравниваемых групп отмечается повышение уровня переваримости при скармливании птице по отношению к контрольной группе и выше на 1,2; 2,9; 3,2 и 2,1 %. Полученные данные показывают, что у птиц опытных групп наблюдается повышенная переваримость компонентов корма и как следствие перепелами усваивается больше питательных веществ, но статистически выраженной разницы между группами не выявлено.
Таким образом, пробиотик Галлобакт-Ф-Плюс стимулирует процесс переваримости компонентов корма, а это в свою очередь способствует наилучшему приросту массы перепелов, особенно опытных групп.
Микробиоценноз химуса кишечника. Микробный фон химуса слепых отростков кишечника исследуемых групп перепелов представлен в таблице 67.
Таблица 67 – Микробный баланс химуса кишечника перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Молочнокислые бактерии, 108 КОЕ/г |
3,4±0,04 | 5,6±0,02* | 7,8±0,05* | 7,9±0,06* | 7,6±0,05* |
| Бифидобактерии, 108 КОЕ/г | 1,6±0,02 | 2,4±0,03* | 3,1±0,03* | 3,1±0,02* | 2,9±0,04* |
| Эшерихии, 102 КОЕ/г | 6,3±0,04 | 5,4±0,04* | 4,0±0,05* | 3,9±0,03* | 4,5±0,04* |
| Энтерококки, 102 КОЕ/г | 5,3±0,02 | 2,1±0,02* | – | – | – |
| Псевдомонады, 10 КОЕ/г | – | – | – | – | – |
| Стафилококки, 102 КОЕ/г | 4,3±0,02 | 3,2±0,02* | 1,4±0,01* | 1,3±0,02* | 1,7±0,01* |
| Стрептококки, 102 КОЕ/г | 5,7±0,03 | 4,1±0,04* | 3,3±0,03* | 3,1±0,03* | 3,6±0,04* |
| Клостридии, 10 КОЕ/г | – | – | – | – | – |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Скармливание перепелам экспериментальных групп комбикорма с пробиотической добавкой Галлобакт-Ф-Плюс оказывает положительное влияние на количественный и качественный состав микробиоты пищеварительного канала. Установлено, что данные полученные при микробиологическом исследовании химуса слепых отростков птицы статистически достоверны. Показано, что титр молочнокислых бактерий в контрольной группе составил 3,4 × 108 КОЕ/г, в то время как в опытных группах был достоверно выше (P < 0,05) в 1,6; 2,2; 2,3
и 2,2 раза. Анализ содержания титра бифидогенных бактерий показал, что количество микроорганизмов было выше в экспериментальных группах, чем
в контрольной, в 1,5; 1,9 и 1,8 раза и является достоверным показателем (P < 0,05). При исследовании химуса установлен рост лакто- и бифидобактерий, при достоверном (P < 0,05) снижении или отсутствии роста и размножения энтеропатогенных бактерий – эшерихий, энтерококков, псевдомонад, клостридий, стафило- и стрептококков, что оказывает положительное влияние на желудочно-кишечный тракт птицы и, как следствие, на результаты выращивания. Так, нами установлено статистически достоверное снижение титра эшерихий
в опытных группах в сравнении с контрольной в 1,2; 1,6 и 1,4 раза (P < 0,05).
В экспериментальных группах выявлено, что количество стафилококков и стрептококков ниже и разница с контролем была достоверна (P < 0,05): в 2,6 раза – для стафилококков и в 1,6 раза – для стрептококков. Также отмечено, что клостридии и псевдомонады не дали роста при их изучении, а уровень энтерококков составил 5,3 × 108 КОЕ/г в контрольной группе, что в 2,5 раза больше к аналогичному показателю в 1-й опытной группе и является достоверным (P < 0,05).
Таким образом, пробиотик Галлобакт-Ф-Плюс способствует стимуляции роста полезной микрофлоры и снижению титра условно-патогенных форм микроорганизмов в кишечнике птиц.
Мясная продуктивность, развитие внутренних органов и качество мяса перепелов. Результаты изучения мясной продуктивности перепелов породы Фараон после применения пробиотика Галлобакт-Ф-Плюс представлены
в таблице 68.
Таблица 68 – Мясная продуктивность перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Живая масса птицы перед убоем, г |
289,78±2,26 | 302,78±2,37 | 310,37±2,08* | 309,67±2,13* | 310,13±2,10* |
| Масса тушки после обескровливания, г | 280,73±2,56 | 293,67±2,68 | 301,32±2,75* | 299,67±2,69* | 300,56±2,73* |
| Масса непотрошеной тушки без перьев, г | 262,89±2,13 | 273,29±2,09 | 281,17±2,12* | 280,67±2,16* | 280,89±2,06* |
| Масса потрошеной тушки, г | 216,84±1,78 | 226,84±2,04 | 232,59±1,86* | 231,58±2,12* | 232,12±2,07* |
| Грудные мышцы, г | 63,49±1,34 | 66,28±1,64 | 69,58±1,43 | 68,90±1,28 | 69,45±1,54 |
| Масса бедренных мышц, г |
22,69±0,89 | 24,16±0,94 | 26,57±0,88 | 26,34±0,82 | 26,47±0,92 |
| Мышцы голени, г | 10,69±0,43 | 11,17±0,46 | 12,67±0,51 | 12,23±0,46 | 12,45±0,49 |
| Остальные мышцы, г | 8,67±0,34 | 9,06±0,27 | 9,87±0,42 | 9,56±0,39 | 9,67±0,40 |
| Всего съедобных мышц, г |
105,54±3,76 | 110,67±3,23 | 118,69±3,65 | 117,03±3,27 | 118,04±3,58 |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
Для установления эффективности влияния изучаемой добавки на мясную продуктивность перепелов проводился контрольный убой. Анализ результатов анатомической разделки птицы показал, что перепела опытных групп по таким показателям мясных качеств, как живая масса птицы перед убоем, масса тушки после обескровливания, а также непотрошеной и потрошеной тушки, достоверно превосходили идентичные показатели в контрольной группе. Так, предубойный вес птицы в контрольной группе составил 289,78 г против 302,78; 310,37; 309,67 и 310,13 г опытных групп, что статистически достоверно выше во 2-й,
3-й и 4-й на 7,1; 6,9 и 7,0 % (Р < 0,05), чем в контроле. Масса тушки после обескровливания, а также масса потрошеной тушки во 2–4-й опытных группах была достоверна выше, чем в контроле в среднем на 7,1 % (Р < 0,05). Тушки птиц экспериментальных групп обладают более высокими мясными качествами – выход съедобных частей в них достигает 95,1; 87,5; 89,1 и 88,1 %, что выше на 4,86; 12,5; 10,9 и 11,9 %, чем у перепелов контрольной группы.
Результаты влияния используемого биопрепарата на массу различных органов и накопление внутриполостного жира птиц представлены в таблице 69.
Таблица 69 – Масса внутренних органов и внутриполосного жира, г (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Легкие | 3,09±0,07 | 3,11±0,10 | 3,11±0,09 | 3,12±0,09 | 3,10±0,07 |
| Печень | 7,37±0,26 | 7,43±0,25 | 7,43±0,31 | 7,42±0,28 | 7,39±0,29 |
| Сердце | 3,32±0,06 | 3,37±0,05 | 3,39±0,07 | 3,40±0,05 | 3,38±0,06 |
| Мышечный желудок | 5,68±0,07 | 5,75±0,11 | 5,76±0,13 | 5,74±0,12 | 5,76±0,09 |
| Железистый желудок | 1,03±0,01 | 1,09±0,02 | 1,11±0,02 | 1,09±0,03 | 1,10±0,02 |
| Кишечник | 9,34±0,23 | 9,45±0,31 | 9,47±0,27 | 9,46±0,29 | 9,48±0,25 |
| Внутриполостной жир | 2,76±0,04 | 2,67±0,05 | 2,57±0,06 | 2,60±0,05 | 2,59±0,08 |
Анализ морфологического состава внутренних органов птицы показывает, что их масса в опытных группах незначительно превышает контрольную, в то время как масса внутриполостного жира имела тенденцию к снижению. Установлено, что статистически достоверной разницы по массе отдельных органов не выявлено, лишь наблюдалось незначительное увеличение в пределах физиологической нормы, что можно объяснить большей массой птицы в экспериментальных группах. Определено, что количество внутриполостного жира у птиц опытных групп было меньше, чем у перепелов контрольной на 3,4; 7,4; 6,2 и 6,6 %, но статистически достоверной разницы не установлено. Данное снижение объясняется тем, что запас внутриполостного жира является весьма мобильным и в первую очередь используется в различных физиологических процессах.
Результаты химического состава мышц перепелов всех групп представлены в таблице 70.
Применение пробиотика Галлобакт-Ф-Плюс не равнозначно сказалось на химическом составе мяса, но достоверной разницы в показателях не выявлено. Так, показатель влаги в ножных и грудных мышцах у птиц контрольной группы составил – 73,49 и 72,36 %, что выше одноименного показателя в опытных группах в среднем на 1,4 и 1,2 %. Содержание белка в ножных мышцах испытуемых групп увеличилось на 1,9; 9,3; 8,2 и 5,7 %, а уровень жира снизился на 2,4; 4,1; 3,6 и 3,3 %. В грудных мышцах соответственно наблюдалось увеличение на 2,9; 6,5; 6,2 и 5,8 % уровня белка и уменьшение на 3,5; 9,1; 5,2 и 6,3 % жира. Определение химического состава мышц перепелов позволило рассчитать индекс качества мяса потрошеных тушек, именно этот показатель характеризует свойства мяса и выражается в отношении «жир : белок», принимая за эталон соотношение 1 : 1. Так, у перепелов опытных групп данный индекс был ниже, чем в контрольной
у ножных мышц – на 5,3; 17,7 и 11,2 %, у грудных мышц – на 8,3 и 18,2 %.
Таблица 70 – Химический состав мышц перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Ножные мышцы | |||||
| Влага, % | 73,49±1,43 | 73,17±1,37 | 71,63±1,38 | 71,85±1,45 | 72,37±1,44 |
| Белок, % | 21,65±0,43 | 22,06±0,41 | 23,67±0,47 | 23,42±0,45 | 22,89±0,51 |
| Жир, % | 4,35±0,11 | 4,25±0,10 | 4,18±0,09 | 4,20±0,08 | 4,21±0,3 |
| Зола, % | 0,51±0,01 | 0,52±0,02 | 0,52±0,02 | 0,53±0,02 | 0,53±0,01 |
| Индекс качества мяса (ИКМ) | 0,20 | 0,19 | 0,17 | 0,18 | 0,18 |
| Грудные мышцы | |||||
| Влага, % | 72,36±1,37 | 71,79±1,29 | 71,06±0,99 | 71,04±1,49 | 71,15±1,22 |
| Белок, % | 23,89±0,69 | 24,57±0,79 | 25,45±0,73 | 25,36±0,77 | 25,28±0,68 |
| Жир, % | 3,23±0,04 | 3,12±0,06 | 2,96±0,08 | 3,07±0,07 | 3,04±0,08 |
| Зола, % | 0,52±0,02 | 0,52±0,01 | 0,53±0,02 | 0,53±0,01 | 0,53±0,02 |
| Индекс качества мяса (ИКМ) | 0,13 | 0,12 | 0,11 | 0,12 | 0,12 |
Таким образом, использование пробиотика Галлобакт-Ф-Плюс способствует повышению диетических особенностей мяса перепелов опытных групп.
Аминокислотный скор мяса перепелов изучали по содержанию незаменимых аминокислот в мышцах птиц (рисунок 30).
Биополноценность мяса определяется его сбалансированным аминокислотным составом, но наиболее объективной оценкой мяса является его аминокислотный скор, а именно содержание в нем незаменимых аминокислот. Анализ данных показывает, что в белке мяса перепелов опытных групп содержалось более оптимальное количество незаменимых аминокислот. Так, уровень лизина в ножных и грудных мышцах опытных групп был выше в среднем на 3,5 %, чем в контрольной. Показатели таких аминокислот как триптофан, фенилаланин, лейцин, метионин имели более высокие показатели в ножных мышцах и были выше в среднем на 3,7; 3,4; 2,7 и 5,3 %, а в грудных мышцах – на 7,7; 2,4; 2,1 и 3,1 %. Таким образом, применение пробиотик Галлобакт-Ф-Плюс в рационе перепелов способствовало повышению биологической полноценности мышечной ткани.
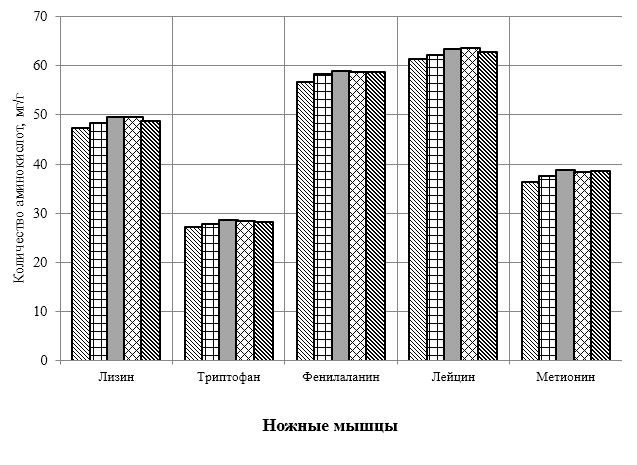
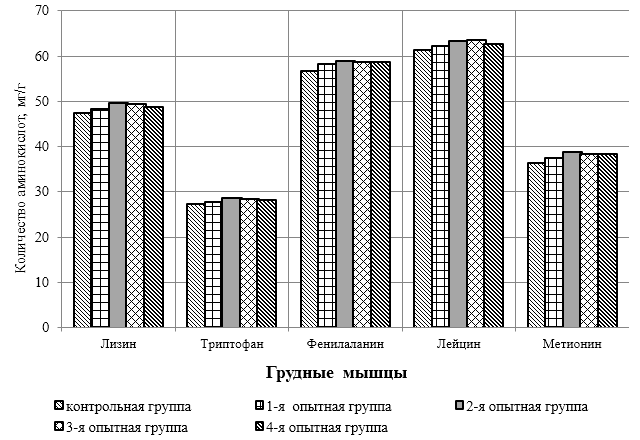
Рисунок 30 – Аминокислотный скор в мышцах перепелов, мг/г
Качество мяса перепелов оценивалось органолептикой, а также проводился дегустационный анализ по пятибалльной шкале (таблица 71).
Таблица 71 − Дегустационная оценка мяса перепелов и бульона из них (n = 10)
| Показатель | Группа | |||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
||
| Ножные мышцы | ||||||
| Аромат | 4,7±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 | |
| Вкус | 4,7±0,2 | 4,8±0,2 | 4,8±0,2 | 4,8±0,2 | 4,8±0,1 | |
| Нежность (жесткость) | 4,7±0,1 | 4,8±0,1 | 4,9±0,1 | 4,9±0,1 | 4,9±0,1 | |
| Сочность | 4,7±0,2 | 4,8±0,1 | 4,9±0,2 | 4,9±0,2 | 4,9±0,2 | |
| Грудные мышцы | ||||||
| Аромат | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | |
| Вкус | 4,8±0,2 | 4,8±0,2 | 4,8±0,2 | 4,8±0,2 | 4,8±0,2 | |
| Нежность (жесткость) |
4,8±0,1 | 4,8±0,1 | 4,9±0,1 | 4,9±0,1 | 4,9±0,2 | |
| Сочность | 4,8±0,2 | 4,8±0,1 | 4,9±0,1 | 4,9±0,2 | 4,9±0,1 | |
| Бульон | ||||||
| Аромат | 4,8±0,1 | 4,8±0,2 | 4,8±0,1 | 4,8±0,1 | 4,8±0,1 | |
| Вкус | 4,8±0,2 | 4,8±0,1 | 4,9±0,2 | 4,9±0,2 | 4,9±0,2 | |
| Прозрачность | 4,8±0,1 | 4,7±0,2 | 4,8±0,1 | 4,8±0,1 | 4,8±0,2 | |
| Крепость (наваристость) |
4,7±0,2 | 4,7±0,1 | 4,8±0,2 | 4,8±0,1 | 4,8±0,2 | |
Органолептический анализ показал, что запах мяса и бульона из него был ароматным, свойственный свежему мясу, посторонних запахов не ощущалось. Результаты дегустационной оценки показали несущественную разницу в разрезе изучаемых групп. Мясо перепелов всех групп имело коричнево-серый цвет, запах свойственный свежему мясу, вкус – приятный. Бульон из-под мяса перепелов опытных и контрольных групп был прозрачный, с незначительным скоплением жира на поверхности. Мясу перепелов и бульону из них в опытных группах дегустационная комиссия дала более высокую оценку, за счет того что оно было более сочным и нежным.
В целом, пробиотик Галлобакт-Ф-Плюс оказывает положительное влияние на мясную продуктивность перепелов породы Фараон, а также способствует улучшению качества получаемой продукции перепеловодства.
Ветеринарно-санитарная экспертиза мяса птицы. При проведении ветеринарно-санитарной оценки обязательным является проведение патологоанатомического вскрытия птицы, чтобы исключить возможные заболевания или патологию на органы и ткани животного вызванную применяемым биопрепаратом. В результате вскрытия перепелов изменений или патологий зафиксировано не было. Органы располагались анатомически правильно, цвет их и окружающих тканей соответствовал цвету здоровой птицы. Посторонних новообразований, кровоизлияний, эрозий, язв во всех группах перепелов – не выявлено.
Согласно ГОСТ Р 54673-2011 тушки перепелов контрольной и опытных групп отнесены к 1-й категории. Так как по показателям упитанности они имели хорошо развитую мускулатуру тела, грудь была округлой формы, наличие подкожного жир – умеренное. Масса переработанной тушки перепелов составляло значение не ниже требований нормативной документации.
Результаты изучения мяса перепелов по физико-химическим и микробиологическим показателям представлены в таблице 72.
Таблица 72 – Физико-химическая и микробиологическая оценка мяса
перепелов (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Р-ция с сернокислой медью | – | – | – | – | – |
| Р-ция с формалином | |||||
| Р-ция на пероксидазу | + | + | + | + | + |
| Кол-во ЛЖК, мг КОН/100 г | 2,65±0,04 | 2,53±0,06 | 2,39±0,04 | 2,40±0,05 | 2,45±0,05 |
| Количество микробных клеток в одном поле зрения микроскопа: | |||||
| – с поверхности тушки | 2,78±0,08 | 2,65±0,09 | 2,64±0,08 | 2,60±0,10 | 3,66±0,07 |
| – с глубоких слоев | – | – | – | – | – |
| рН мяса, ед. | |||||
| Первые сутки | 6,83±0,17 | 6,81±0,20 | 6,76±0,19 | 6,79±0,19 | 6,81±0,18 |
| Вторые сутки | 6,56±0,18 | 6,48±0,18 | 6,45±0,21 | 6,39±0,15 | 6,43±0,20 |
| Третьи сутки | 6,08±0,15 | 6,03±0,21 | 5,98±0,18 | 6,07±0,22 | 5,97±0,19 |
По результатам ветеринарно-санитарной экспертизы установлено, что мясо, полученное от птицы, соответствует нормативной документации. Так, при проведении реакции с сернокислой медью и формалином показало отрицательный результат, на пероксидазу – положительный, что говорит о том, что мясо получено от здоровой птицы. При изучении летучих жирных кислот в мясе перепелов установлено, что их количество во всех группах было в пределах допустимых требований и составило в контрольной – 2,65 мг КОН/100 г, против 22,65; 2,64; 2,60 и 3,66 мг КОН/100 г в 1-й, 2-й, 3-й и 4-й опытных группах. Изучение количества микробных клеток с поверхности тушки и с ее глубоких слоев показало, что показатели находятся в пределах допустимых норм. Исследование кислотности мяса выявило резкое снижение показателя рН мышц в течение первых трех суток, которое характерно для мяса, полученного от здоровой птицы. Следовательно, применение пробиотика Галлобакт-Ф-Плюс не оказало негативного воздействия на тушки перепелов и их мясо, полученные результаты показали соответствие, изучаемых параметров нормативной документации, а полученные продукты допускаются к употреблению.
Таким образом, результаты проведенных исследований показали, что пробиотик Галлобакт-Ф-Плюс обладает стимулирующим действием на обмен веществ, повышает приросты, сохранность, способствует высоким показателям биополноценностии мясной продукции перепеловодства. Мясо перепелов может быть использовано в пищу независимо от дозы и сроков использования испытуемого биопрепарата.
В целом, результаты изучения фармакологических свойств пробиотика Галлобакт-Ф-Плюс, а также подбор эффективной его дозы показали, что по совокупности эффекта наилучшей дозой для перепелов можно считать 0,5 мл/гол. Данное количество пробиотика в рационе перепелов породы Фараон позволяет проявить высокий биоресурстный потенциал птицы.
Экономическая эффективность использования пробиотика. Результаты экономической эффективности использования пробиотика Галлобакт-Ф-Плюс на перепелах породы Фараон представлены в таблице 73.
Таблица 73 – Экономическая эффективность использования пробиотика Галлобакт-Ф-Плюс
| Показатель | Группа | |||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Поголовье в начале опыта, гол. | 200 | 200 | 200 | 200 | 200 | |
| Сохранность, % | 93,5 | 95,5 | 98,5 | 98,5 | 97,5 | |
| Поголовье в конце опыта, гол. | 187 | 191 | 197 | 197 | 195 | |
| Масса тушки на реализацию | ||||||
| Масса потрошенной тушки, г | 216,84±1,78 | 226,84±2,04 | 232,59±1,86* | 231,58±2,12* | 232,12±2,07* | |
| Масса потрошенной тушки всего, кг | 40,55 | 43,32 | 45,82 | 45,62 | 45,26 | |
| Затраты комбикорма за период выращивания | ||||||
| На голову, г | 937,85 | 980,37 | 990,40 | 993,65 | 989,49 | |
| На все поголовье, кг | 175,38 | 187,25 | 195,11 | 195,75 | 192,95 | |
| Затраты пробиотика | ||||||
| Всего, мл | – | 477,5 | 985,0 | 1 477,5 | 1950,0 | |
| Экономическая эффективность | ||||||
| Цена 1 кг комбикорма, руб. | 30 | |||||
| Стоимость израсходованного корма всего, руб. |
5 261,40 | 5 617,50 | 5 853,30 | 5 872,5 | 5 788,5 | |
| Цена 1 л пробиотика, руб. | – | 60 | ||||
| Стоимость пробиотика всего, руб. | – | 28,65 | 59,10 | 88,65 | 117,00 | |
| Стоимость израсходованного корма и пробиотика, руб. |
5 261,40 | 5 646,15 | 5 912,40 | 5 961,15 | 5 905,50 | |
| Цена реализации 1 кг мяса перепелов, руб. | 400,0 | |||||
| Выручка от реализации мяса птицы, руб. | 16 220,00 | 17 328,00 | 18 328,00 | 18 248,00 | 18 104,00 | |
| Прибыль от реализации мяса птицы, руб. | 10 958,60 | 11 681,85 | 12 415,60 | 12 286,85 | 12 198,50 | |
| Экономический эффект от использования пробиотика к контролю,
руб. % |
–
– |
723,25
6,60 |
1 457,00
13,30 |
1 328,25
12,12 |
1 239,90
11,31 |
|
В результате проведенного предварительного анализа экономической эффективности применения в рационе перепелов породы Фараон пробиотической добавки Галлобакт-Ф-Плюс выявлено, что с учетом затрат на комбикорм
и микробной добавки, прибыль в 1–4-й опытных группах составила 11 681,85; 12 415,60; 12 286,85 и 12 198,50 руб., против 10 958,60 руб. в контрольной группе, что выше на 723,25; 1 457,00; 1 328,25 и 1 239,90 руб., соответственно. В целом, наилучшие результаты были получены во 2-й опытной группе, в которой экономический эффект превышал контрольную группу на 13,30 %.
Таким образом, экономически целесообразно проводить производственную апробацию пробиотика Галлобакт-Ф-Плюс на перепелах породы Фараон
в дозе, составляющей 0,5 мл/гол.
2.16.3 Фармакологическое действие пробиотика Галлобакт-Ф-Плюс на организм цыплят-бройлеров быстрорастущих кроссов
Следующим этапом исследований явилось изучить влияние пробиотической добавки Галлобакт-Ф-Плюс на организм цыплят-бройлеров быстрорастущих кроссов. В качестве объектов исследований использовали цыплят-бройлеров кроссов Росс 308 и Кобб 500.
Влияние пробиотика Галлобакт-Ф-Плюс на цыплят-бройлеров
кросса Росс 308
Для изучения влияния пробиотика Галлобакт-Ф-Плюс на сохранность, продуктивность, основные показатели обмена веществ, а также качество продукции от цыплят-бройлеров кросса Росс 308 был проведен научно-хозяйственный опыт, в котором методом групп-аналогов было сформировано пять групп птиц по 60 гол. в каждой: контрольная группа – птица получала стандартный рацион, рекомендованный ВНИТИП, и имела свободный доступ к воде; первая опытная группа – подопытная птица получала стандартный рацион,
а также дополнительно в воду вводился пробиотик Галлобакт-Ф-Плюс в дозе 0,25 мл/гол.; вторая опытная группа – Галлобакт-Ф-Плюс вводили в дозе 0,5 мл/гол.; третья опытная группа – объекты исследований получали стандартный рацион, рекомендованный ВНИТИП, а также в воду добавляли пробиотик в дозе 0,75 мл/гол.; четвертая опытная группа – объекты исследований получали стандартный рацион, а в воду добавляли пробиотик в дозе 1,0 мл/гол. Опыт продолжался 42 дня.
Рост и развитие цыплят-бройлеров. Для проведения научно-хозяйственного опыта на первые сутки рождения формировали группы из клинически здоровых цыплят, живая масса которых была практически аналогичной, с учетом требований. В процессе выращивания птицы наблюдали за ее поведением, перьевым покровом, поеданием корма. Установлено, что изучаемые клинические показатели были удовлетворительными. Данные динамики живой массы, прироста, употребленного комбикорма птицей, а также сохранности представлены в таблице 74 и на рисунке 31.
Таблица 74 – Изучаемые хозяйственные показатели цыплят-бройлеров
кросса Росс 308, (n = 60)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Сохранность, % | 93,3 | 96,6 | 96,6 | 100 | 100 |
| Прирост живой массы за период выращивания (042 сут) | |||||
| Одной головы в среднем, г |
2636,18 | 2661,37 | 2682,37 | 2712,92 | 2709,42 |
| Расход комбикорма за период выращивания (042 сут) | |||||
| Одной головы, г | 4798,74 | 4814,25 | 4828,76 | 4836,41 | 4843,68 |
| Конверсия, кг | 1,82 | 1,81 | 1,80 | 1,78 | 1,79 |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
Результаты изучения хозяйственных показателей при выращивании цыплят-бройлеров кросса Росс 308 показали, что применение пробиотика Галлобакт-Ф-Плюс способствовало повышению сохранности птицы 1-й и 2-й опытных групп по сравнению с контрольной на 3,3 %, а в 3-й и 4-й опытных группах данный показатель был максимальным и выше, чем в контрольной, на 6,7 %.
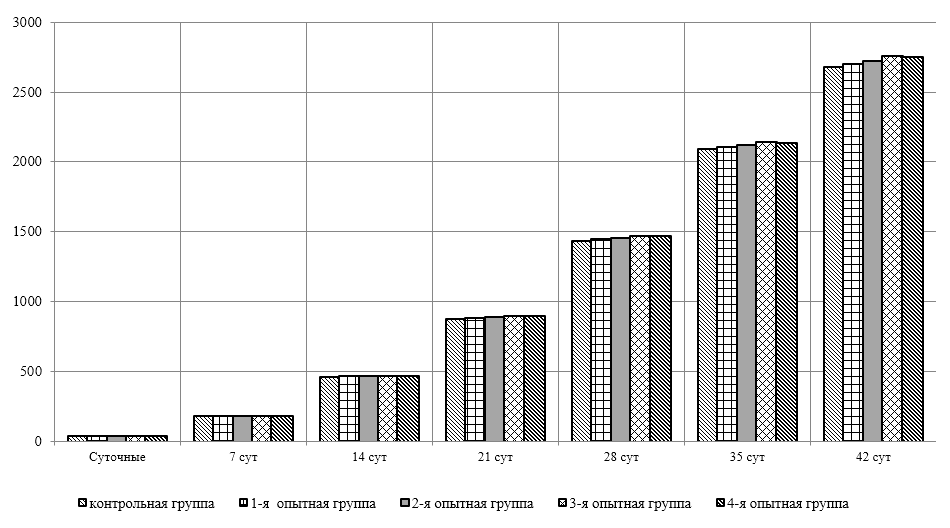
Рисунок 31 ‒ Динамика живой массы, г
Изучение живой массы птиц показало, что в опытных группах изучаемый показатель в каждый период взвешивания цыплят-бройлеров равномерно повышался в зависимости от дозы пробиотика. Однако статистически достоверная разница была выявлена на 35-е сутки выращивания птицы в 3-й и 4-й опытных группах по сравнению с контрольной, в которых масса оказалась выше на 2,4
и 2,3 % (P < 0,05). В 1-й и 2-й опытных группах также масса птиц была выше, чем в контрольной, на 0,9 и 1,3 %, но разница была не достоверна. На 42-й день взвешивания масса цыплят-бройлеров во 2-й, 3-й и 4-й опытных группах была выше, чем в контрольной, на 1,7; 2,8 и 2,7 % при статистически достоверной разнице (P < 0,05). В целом, прирост цыплят-бройлеров в контрольной группе составил 2636,18 г, а в 1–4-й опытных группах – 2661,37; 2682,37; 2712,92 и 2709,42 г, соответственно, что выше, чем в контроле на 1,0; 1,6; 2,9 и 2,8 %.
В связи с тем, что масса птиц опытных групп была выше, чем в контрольной, у них наблюдалась повышенная поедаемость комбикормов. Однако анализ конверсии комбикорма показал, что изучаемый показатель в опытных группах был ниже, чем в контрольной на 0,01; 0,02; 0,04 и 0,03 кг.
Таким образом, применение биопрепарата на основе живых культур лактобацилл способствовал повышению сохранности, живой массы цыплят-бройлеров,
а также снижению показателя конверсии корма, что максимально было выражено в 3-й опытной группе при дозе пробиотика Галлобакт-Ф-Плюс 0,75 мл/гол.
Морфологические и биохимические показатели крови. Показатели цельной
и сыворотки крови являются главнейшими интерьерными значениями состояния организма птицы, в особенности при использовании в их рационе новых биопрепаратов. Количественный и качественный состав крови говорит об интенсивности обменных процессов, а также связанных с ними процессах развития и продуктивности птицы. Результаты морфологических и биохимических показателей крови цыплят-бройлеров кросса Росс 308 представлены в таблице 75.
Результаты анализа цельной крови и ее сыворотки на 21-е сутки исследований не дали статистически достоверных изменений в показателях и в разрезе изучаемых подопытных групп все они находились в пределах физиологической нормы (таблица 75). Однако изучение данных показателей на 42-е сутки выращивания цыплят-бройлеров имели не только тенденцию к их улучшению, но
и в отдельных опытных группах были достоверно изменены по сравнению
с контрольной. Так, в 3-й и 4-й опытных группах были зафиксированы статистические различия по ряду показателей, в частности, по уровню эритроцитов, которых в данных группах было выше, чем в контрольной на 8,1 и 7,3 %, гемоглобина – на 4,7 и 4,6 %, общего белка – на 4,6 %, альбуминов – на 12,9
и 11,1 %, кальция – на 4,8 и 4,6 %, БАСК – на 4,5 и 4,1 %, ЛАСК – на 3,6
и 3,4 % (Р < 0,05). Также в 3-й и 4-й опытных группах было выявлено достоверное снижение уровня холестерина в сыворотки крови цыплят-бройлеров кросса Росс 308 по сравнению с контрольной на 9,3 и 8,5 % (Р < 0,05).
Таблица 75 – Морфо-биохимический статус крови цыплят-бройлеров
кросса Росс 308
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| 1 | 2 | 3 | 4 | 5 | 6 |
| 21-е сутки | |||||
| Эритроциты, 1012/л | 2,64±0,04 | 2,65±0,03 | 2,69±0,03 | 2,67±0,05 | 2,70±0,03 |
| Гемоглобин, г/л | 83,34±1,45 | 83,45±1,54 | 83,54±1,60 | 84,12±1,63 | 83,89±1,62 |
| Тромбоциты, 109/л | 62,34±0,86 | 62,51±0,76 | 62,79±0,71 | 62,97±0,64 | 62,76±0,72 |
| Лейкоциты, 109/л | 23,34±0,41 | 23,44±0,35 | 23,49±0,39 | 23,38±0,33 | 23,43±0,30 |
| Общий белок, г/л | 36,76±0,54 | 36,79±0,53 | 36,87±0,50 | 37,12±0,43 | 37,05±0,47 |
|
14,54±0,22 | 14,65±0,15 | 14,79±0,18 | 15,11±0,16 | 14,97±0,20 |
|
22,22±0,36 | 22,14±0,28 | 22,08±0,33 | 22,01±0,30 | 22,08±0,26 |
| А/Г коэффициент | 0,65±0,01 | 0,66±0,02 | 0,67±0,01 | 0,68±0,02 | 0,67±0,03 |
| Холестерин, мМ/л | 3,54±0,06 | 3,56±0,05 | 3,53±0,04 | 3,51±0,04 | 3,53±0,05 |
| АСТ, Ед/л | 213,36±2,74 | 214,84±2,46 | 214,47±2,57 | 213,45±2,54 | 213,54±2,50 |
| АЛТ, Ед/л | 22,65±0,26 | 22,73±0,29 | 22,84±0,22 | 22,79±0,19 | 22,64±0,24 |
| Кальций, мМ/л | 9,58±0,11 | 9,59±0,09 | 9,63±0,12 | 9,64±0,10 | 9,62±0,14 |
| Фосфор, мМ/л | 4,57±0,05 | 4,59±0,08 | 4,62±0,06 | 4,63±0,07 | 4,65±0,08 |
| БАСК, % | 37,8±1,87 | 38,1±1,82 | 38,2±1,73 | 38,3±1,66 | 38,2±1,78 |
| ЛАСК, % | 32,5±1,24 | 32,7±1,19 | 32,9±1,29 | 32,9±1,18 | 32,7±1,22 |
| 42-е сутки | |||||
| Эритроциты, 1012/л | 2,73±0,02 | 2,76±0,01 | 2,80±0,03 | 2,95±0,02* | 2,93±0,03* |
| Гемоглобин, г/л | 84,35±0,53 | 84,78±0,58 | 84,98±0,61 | 88,34±0,60* | 88,25±0,52* |
| Тромбоциты, 109/л | 64,54±0,74 | 64,87±0,71 | 66,21±0,62 | 66,57±0,68 | 66,52±0,70 |
| Лейкоциты, 109/л | 24,56±0,34 | 24,65±0,31 | 24,62±0,33 | 24,68±0,30 | 23,61±0,31 |
| Общий белок, г/л | 37,45±0,21 | 37,76±0,28 | 38,12±0,24 | 39,23±0,26* | 39,18±0,27* |
|
14,78±0,25 | 14,86±0,27 | 15,56±0,29 | 16,68±0,27* | 16,42±0,29* |
|
22,67±0,31 | 22,90±0,29 | 22,56±0,35 | 22,55±0,27 | 22,76±0,30 |
| Продолжение таблицы 75 | |||||
| 1 | 2 | 3 | 4 | 5 | 6 |
| А/Г коэффициент | 0,65±0,03 | 0,65±0,02 | 0,68±0,03 | 0,73±0,01 | 0,72±0,02 |
| Холестерин, мМ/л | 3,78±0,02 | 3,75±0,04 | 3,62±0,06 | 3,43±0,03* | 3,46±0,02* |
| АСТ, Ед/л | 227,45±2,54 | 225,41±2,49 | 225,32±2,47 | 224,67±2,64 | 224,87±2,56 |
| АЛТ, Ед/л | 23,45±0,28 | 23,33±0,32 | 23,25±0,31 | 23,15±0,29 | 23,21±0,25 |
| Кальций, мМ/л | 9,34±0,05 | 9,43±0,09 | 9,54±0,06 | 9,79±0,05* | 9,77±0,06* |
| Фосфор, мМ/л | 4,21±0,07 | 4,32±0,08 | 4,37±0,07 | 4,45±0,08 | 4,44±0,07 |
| БАСК, % | 41,0±0,25 | 41,7±0,32 | 42,3±0,28 | 45,5±0,30* | 45,12±0,27* |
| ЛАСК, % | 32,8±0,28 | 33,4±0,24 | 33,8±0,22 | 36,4±0,21* | 36,2±0,27* |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
Таким образом, результаты анализа крови цыплят-бройлеров кросса Росс 308 свидетельствовали о том, что на конец эксперимента у птиц опытных групп, в рационе которых использовали пробиотик Галлобакт-Ф-Плюс, наблюдалась активизация, а также стимуляция обменных процессов, с одновременным усилением иммунного статуса птицы за счет повышения показателей бактерицидной
и лизоцимной активностей сыворотки крови, что в итоге обеспечило высокий показатель сохранности и продуктивности сельскохозяйственной птицы.
Влияние пробиотика на пищеварение. В конце научно-хозяйственного эксперимента проводили балансовый опыт для изучения показателей переваримости компонентов комбикорма. Данные представлены на рисунке 32.
Установлено, что показатели переваримости изучаемых питательных веществ комбикорма в опытных группах были незначительно выше, чем в контрольной. Так, переваримость органического вещества комбикорма в 1–4-й опытных группах составили 80,78; 81,67; 82,47 и 82,32 %, против 80,04 %
в контрольной группе (разница – 0,74; 1,63; 2,43 и 2,28 %, соответственно). Переваримость сырого жира также имела тенденцию к повышению в опытных группах по сравнению с контрольной на 0,31; 0,56; 1,4 и 1,1 %, клетчатки – на 0,32; 0,6; 0,95 и 0,87 %, БЭВ – на 0,73; 1,2; 1,61 и 1,4 %, соответственно. По переваримости сырого протеина в разрезе исследуемых групп была выявлена
в 3-й и 4-й опытных группах статистически достоверная разница по сравнению с контрольной и составила 79,51 и 78,79 % против 74,54 %, что соответственно выше, чем в контроле на 5,0 и 4,3 % (Р < 0,05).
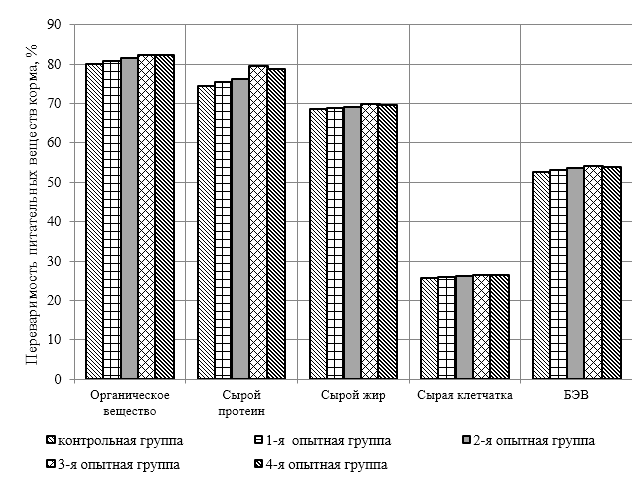
Рисунок 32 – Переваримость питательных веществ комбикорма
цыплятами-бройлерами, %
Таким образом, применение в составе рациона птиц пробиотика Галлобакт-Ф-Плюс поспособствовало лучшему перевариванию веществ комбикорма, что обеспечило повышение продуктивности цыплят-бройлеров опытных групп.
Микробиоценноз химуса кишечника. Для изучения влияния испытуемого биопрепарата Галлобакт-Ф-Плюс на формирование микробного фона в слепых отростках кишечника цыплят-бройлеров была проведена посмертная резекция данного отдела желудочно-кишечного тракта с последующим микробиологическим анализом. Результаты микробиологических исследований слепых отростков кишечника птиц представлены в таблице 76.
Микробиоценноз кишечника птиц всех опытных групп по уровню нормофлоры достоверно превосходил данный показатель в контрольной группе. По содержанию молочнокислых бактерий ее титр в 1–4-й опытных группах был выше, чем в контрольной в 1,4; 1,6; 1,9 и 1,8 раз (P < 0,05). Уровень бифидобактерий достоверно был повышен в опытных группах по сравнению с контролем в 1,1; 1,2
и 1,3 раза (P < 0,05). Также в опытных группах была выявлена статистически достоверное снижение уровня условно-патогенной микрофлоры, а по отдельным группам изучаемой микрофлоры наблюдалось ее отсутствие в кишечнике цыплят-бройлеров. Так, содержание бактерий группы кишечной палочки в контрольной группе составило 6,7 × 103 КОЕ/г, против 4,4 × 103 КОЕ/г в 1-й и 2-й опытных группах; 2,8 и 3,1 × 103 КОЕ/г в 3-й и 4-й опытных группах (P < 0,05). Наблюдалось достоверное снижение стафилококков и стрептококков в опытных группах по сравнению с контролем в 1,2; 1,4; 1,9 и 1,8 раза, а также 1,2; 1,3; 1,5 и 1,4 раза, соответственно (P < 0,05). В 1-й опытной группе было зафиксировано достоверное снижение по сравнению с контрольной группой содержание энтерококков
в 3,2 раза (P < 0,05). При этом во 2-й, 3-й и 4-й опытных группах не было выявлено данной группы микроорганизмов. Псевдомонады в контрольной группе составили 3,2 × 10 КОЕ/г, а титр клостридий – 2,5 × 10 КОЕ/г, в то время как в 1–4-й опытных группах данную микрофлору не выявили.
Таблица 76 – Баланс полезной и условно-патогенной микрофлоры химуса
слепых отростков цыплят-бройлеров (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Молочнокислые бактерии, 108 КОЕ/г |
2,3±0,05 | 3,2±0,04* | 3,8±0,03* | 4,4±0,03* | 4,3±0,06* |
| Бифидобактерии, 108 КОЕ/г | 3,2±0,07 | 3,5±0,05* | 3,8±0,05* | 4,1±0,06* | 4,1±0,04* |
| Эшерихии, 103 КОЕ/г | 6,7±0,07 | 4,4±0,06* | 3,7±0,05* | 2,8±0,04* | 3,1±0,05* |
| Энтерококки, 102 КОЕ/г | 6,8±0,04 | 2,1±0,05* | – | – | – |
| Псевдомонады, 10 КОЕ/г | 3,2±0,03 | – | – | – | – |
| Стафилококки, 102 КОЕ/г | 6,4±0,04 | 5,3±0,03* | 4,5±0,04* | 3,3±0,04* | 3,5±0,03* |
| Стрептококки, 102 КОЕ/г | 5,5±0,05 | 4,7±0,04* | 4,2±0,05* | 3,7±0,04* | 3,8±0,03* |
| Клостридии, 10 КОЕ/г | 2,5±0,05 | – | – | – | – |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Таким образом, пробиотик Галлобакт-Ф-Плюс за счет наличия в своем составе полезных лактобактерий, обладающих высокими пробиотическими свойствами, способствовал угнетению развития патогенной микробиоты, при этом оказывал позитивное влияние на рост нормофлоры кишечника.
Мясная продуктивность, развитие внутренних органов и качество мяса птиц. Показателем продуктивности ссельскохозяйственных животных, в том числе птицы, являются не только данные прироста, но и доля нарощенной мышечной ткани. Данные убойного выхода цыплят-бройлеров кросса Росс 308 представлены в таблице 77.
Таблица 77 – Убойные показатели цыплят-бройлеров (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Предубойная масса птицы, г | 2658,76±10,56 | 2697,57±10,25 | 2711,67±10,87 | 2747,73±10,37* | 2743,26±10,43* |
| Масса потрошеной тушки, г | 1813,27±10,56 | 1853,23±10,87 | 1873,76±11,04 | 1912,42±11,35* | 1903,82±11,42* |
| Убойный выход, % | 68,2 | 68,7 | 69,1 | 69,6 | 69,4 |
Изучая убойные показатели цыплят-бройлеров кросса Росс 308 при использовании в их рационе пробиотика Галлобакт-Ф-Плюс было установлено, что биопрепарат в 3-й и 4-й опытных группах статистически достоверно повысил массу потрошенной тушки по сравнению с контрольной группой на 5,5
и 5,0 % (P < 0,05). Результаты убойного выхода в 1–4-й опытных группах составили 68,7; 69,1; 69,6 и 69,4 %, что выше, чем в контрольной, на 0,5; 0,9; 1,4
и 1,2 %, соответственно.
Такие показатели тушки как ее морфологический состав, а именно соотношение мышечной, кожной и костной тканей друг к другу, определяют количественные и качественные особенности мясной продуктивности цыплят-бройлеров. В связи с чем, проводился морфологический состав грудных и ножных мышц тушки подопытных птиц. Результаты разделки грудины на части представлены в таблице 78.
Анализ морфологического состава грудки цыплят-бройлеров кросса Росс 308 показал, что масса грудных мышц в опытных группах была выше, чем
в контрольной на 12,54; 20,86; 39,21 и 31,44 г. Доля данной группы мышц
в контроле по отношению к массе потрошенной тушки составила 22,1 %, в то время как в 1–4-й опытных группах этот показатель был выше на 0,2; 0,4; 0,8
и 0,6 %. Масса кожи и костей грудной части в опытных группах также была незначительно выше, чем в контрольной, что связано с большей массой тела птицы. В целом, масса всех тканей груди в опытных группах была выше, чем
в контрольной на 2,9; 5,5; 10,2 и 8,6 %, а на долю их пришлось 29,3; 29,7; 30,4
и 30,1 % против 29,1 % в контрольной группе.
Данные по морфологическому составу бедренной части цыплят-бройлеров представлены в таблице 79.
При изучении морфологического состава бедра цыплят-бройлеров установлено, что выход бедренных мышц без кости и кожи во 2-й, 3-й и 4-й опытных группах был выше, чем в контрольной, на 0,1; 0,5 и 0,3 %, а их масса превосходила – на 4,2; 9,8 и 7,6 %, соответственно. В 1-й опытной группе выход бедренных мышц по отношению к массе потрошенной тушки был идентичен контрольной, однако масса изучаемых мышц была выше на 2,2 %. Масса кожи бедренной части 1–4-й опытных групп незначительно превосходила изучаемый показатель в контрольной группе на 0,88; 3,2; 6,0 и 5,8 г, но выход данного показателя был практически на одном уровне и составил в контроле 2,2 %, а в опытных – 2,2; 2,3; 2,4 и 2,4 %, соответственно. В целом, по массе всех частей бедра максимальное значение было зафиксировано в 3-й опытной группе, которое составило 338,52 г, в 4-й опытной группе – 333,19 г, во 2-й группе – 322,33 г и 1-й опытной группе – 313,21 г, против 306,50 г в контрольной группе.
Морфологический состав голени птиц представлен в таблице 80.
Аналогичная тенденция в пользу опытных групп была выявлена при анализе морфологического состава голени цыплят-бройлеров кросса Росс 308 при использовании пробиотика Галлобакт-Ф-Плюс. Масса всех тканей голени (мышцы, кожа и кости) в 1–4-й опытных группах превосходила контрольную группу на 2,2; 5,5; 7,7 и 7,2 %, соответственно, а выход данной части тела птиц по отношению к потрошенной тушки во 2–4-й опытных группах был выше на 0,5 и 0,3 %.
Данные по состоянию и развитию органов цыплят-бройлеров кросса Росс 308 представлены в таблице 81.
Таблица 78 – Морфологический состав грудки цыплят-бройлеров (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Масса потрошеной тушки | г | 1813,27±10,56 | 1853,23±10,87 | 1873,76±11,04 | 1912,42±11,35* | 1903,82±11,42* |
| Масса грудных мышц | г | 400,73±5,87 | 413,27±5,69 | 421,59±5,82 | 439,94±5,76 | 432,17±5,75 |
| % | 22,1 | 22,3 | 22,5 | 22,9 | 22,7 | |
| Масса кожи | г | 56,21±1,12 | 57,45±1,13 | 59,96±1,22 | 63,11±1,04 | 62,83±1,09 |
| % | 3,1 | 3,1 | 3,2 | 3,3 | 3,3 | |
| Масса костей | г | 70,72±2,32 | 72,28±2,43 | 74,95±2,35 | 78,41±2,23 | 78,06±2,21 |
| % | 3,9 | 3,9 | 4,0 | 4,1 | 4,1 | |
| Масса всех тканей | г | 527,66±6,75 | 543,00±6,54 | 556,50±6,86 | 581,46±6,59 | 573,06±6,64 |
| % | 29,1 | 29,3 | 29,7 | 30,4 | 30,1 | |
Таблица 79 – Морфологический состав бедра птицы (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Масса потрошеной тушки | г | 1813,27±10,56 | 1853,23±10,87 | 1873,76±11,04 | 1912,42±11,35* | 1903,82±11,42* |
| Масса бедренных мышц | г | 223,04±3,45 | 227,95±3,25 | 232,35±3,47 | 244,79±3,44 | 239,89±3,36 |
| % | 12,3 | 12,3 | 12,4 | 12,8 | 12,6 | |
| Масса кожи | г | 39,90±0,78 | 40,78±0,67 | 43,10±0,72 | 45,90±0,58 | 45,70±0,65 |
| % | 2,2 | 2,2 | 2,3 | 2,4 | 2,4 | |
| Масса костей | г | 43,52±0,87 | 44,48±0,83 | 46,85±0,76 | 47,81±1,11 | 47,60±0,95 |
| % | 2,4 | 2,4 | 2,5 | 2,5 | 2,5 | |
| Масса всех тканей | г | 306,5±5,87 | 313,2±5,89 | 322,3±6,23 | 338,5±6,21 | 333,19±5,98 |
| % | 16,9 | 16,9 | 17,2 | 17,7 | 17,5 | |
Таблица 80 – Морфологический состав голени птицы (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Масса потрошеной тушки | г | 1813,27±10,56 | 1853,23±10,87 | 1873,76±11,04 | 1912,42±11,35* | 1903,82±11,42* |
| Масса мышц голени | г | 157,76±2,45 | 161,24±2,35 | 164,89±2,54 | 168,30±2,46 | 167,54±2,49 |
| % | 8,7 | 8,7 | 8,8 | 8,8 | 8,8 | |
| Масса кожи | г | 38,08±0,87 | 38,92±0,95 | 41,23±0,82 | 42,08±0,88 | 41,89±0,82 |
| % | 2,1 | 2,1 | 2,2 | 2,2 | 2,2 | |
| Масса костей | г | 59,84±0,81 | 61,16±0,76 | 63,71±0,80 | 65,03±0,80 | 64,73±0,94 |
| % | 3,3 | 3,3 | 3,4 | 3,4 | 3,4 | |
| Масса всех тканей | г | 255,68±4,56 | 261,32±4,87 | 269,83±4,86 | 275,41±4,68 | 274,16±4,69 |
| % | 14,1 | 14,1 | 14,6 | 14,4 | 14,4 | |
Таблица 81 – Развитие внутренних органов цыплят-бройлеров (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Предубойная масса птицы | г | 2658,76±10,56 | 2697,57±10,25 | 2711,67±10,87 | 2747,73±10,37* | 2743,26±10,43* |
| Печень | г | 36,64±0,62 | 36,85±0,73 | 37,26±0,74 | 37,38±0,87 | 37,32±0,93 |
| % | 1,38 | 1,37 | 1,38 | 1,36 | 1,36 | |
| Сердце | г | 11,74±0,32 | 11,86±0,44 | 12,10±0,55 | 12,22±0,46 | 12,15±0,31 |
| % | 0,45 | 0,44 | 0,45 | 0,44 | 0,44 | |
| Мышечный желудок | г | 31,31±0,82 | 31,53±0,64 | 31,95±0,57 | 32,33±0,78 | 32,18±0,63 |
| % | 1,18 | 1,17 | 1,18 | 1,18 | 1,17 | |
| Железистый желудок | г | 10,10±0,33 | 10,37±0,34 | 10,74±0,45 | 11,03±0,42 | 10,98±0,37 |
| % | 0,38 | 0,38 | 0,40 | 0,40 | 0,40 | |
| Селезенка | г | 4,03±0,11 | 4,11±0,16 | 4,13±0,13 | 4,17±0,09 | 4,15±0,10 |
| % | 0,16 | 0,15 | 0,15 | 0,15 | 0,15 | |
| Кишечник | г | 117,76±1,13 | 118,65±1,10 | 119,34±1,17 | 120,79±1,20 | 120,16±1,15 |
| % | 4,43 | 4,39 | 4,40 | 4,40 | 4,38 | |
| Легкие | г | 10,54±0,36 | 10,65±0,32 | 10,75±0,26 | 11,12±0,28 | 10,97±0,33 |
| % | 0,40 | 0,39 | 0,40 | 0,40 | 0,40 | |
| Почки | г | 8,76±0,26 | 8,79±0,21 | 8,83±0,27 | 8,86±0,23 | 8,79±0,26 |
| % | 0,33 | 0,33 | 0,33 | 0,32 | 0,32 | |
| Желчный пузырь | г | 2,45±0,07 | 2,48±0,06 | 2,49±0,08 | 2,52±0,05 | 2,51±0,07 |
| % | 0,1 | 0,09 | 0,09 | 0,09 | 0,09 | |
Результаты анализа развития (массы) представленных в таблице 81 внутренних органов показали, что в разрезе исследуемых групп статистически достоверной разницы не наблюдалось. Расположение органов было анатомически правильным, размер и консистенция их свидетельствовали об отсутствии той или иной патологии. Соотношение исследуемых органов в разрезе групп к массе тела птицы были почти идентичны.
Незначительная разница в массе органов между контрольной и опытными группами зависела от превосходившей в опытных группах массы тела и не связана с патологическими процессами. Так, например, масса печени в опытных группах была больше, чем в контрольной на 0,6; 1,7; 2,0 и 1,9 %, соответственно; масса сердца – на 1,0; 3,0; 4,1 и 3,5 % и т. д. Было изучено влияние пробиотика на развитие эндокринных желез у цыплят-бройлеров кросса Росс 308, данные которых представлены в таблице 82. Эндокринные железы продуцируют гормоны, которые из клеток железы попадают в кровяное русло. В свою очередь гормоны, разносятся с током крови по всему организму, тем самым регулируя все основные процессы жизнедеятельности (Фисинин В. И. [и др.], 2016).
Таблица 82 – Развитие эндокринных желез цыплят-бройлеров, мг (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Щитовидная железа |
183,56±1,45 | 184,21±1,49 | 184,35±1,57 | 184,43±1,55 | 184,41±1,41 |
| Паращитовидная железа | 15,23±0,45 | 15,36±0,41 | 15,58±0,47 | 15,87±0,35 | 15,75±0,44 |
| Надпочечник | 165,86±1,87 | 166,12±1,89 | 166,32±1,85 | 166,83±1,76 | 166,73±1,74 |
| Фабрициева сумка |
2,67±0,04 | 2,76±0,05 | 2,81±0,07 | 2,87±0,05 | 2,88±0,04 |
| Поджелудочная железа | 2,85±0,03 | 2,89±0,06 | 2,95±0,06 | 2,99±0,04 | 2,97±0,05 |
Результаты исследований показали, что применение пробиотика Галлобакт-Ф-Плюс в опытных группах способствовало незначительному изменению массы изучаемых желез в сторону их увеличения по сравнению с контрольной группой, но без достоверной разницы. Так, масса щитовидной железы, отвечающая за выработку гормона тироксина, который повышает уровень окислительных процессов в тканях, обладает катаболическим действием, стимулируя процессы диссимиляции, ускоряет дифференцировку тканей и органов, способствует смене оперения сельскохозяйственной птицы, в 1-й и 2-й опытных группах была больше, чем в контрольной, на 0,4 %, а в 3-й и 4-й – на 0,5 %. Масса паращитовидной железы, принимающей косвенное участие в укреплении костного скелета благодаря вырабатываемому гормону, который участвует в регуляции обмена кальция, в 1-й опытной группе была больше, чем в контрольной на 0,8 %, во 2-й опытной группе – на 2,3 %, в 3-й опытной группе – на 4,2 %, а в 4-й группе – на 3,4 %. Большое значение в организме имеют надпочечники, масса которых в опытных группах также была незначительно больше, чем
в контрольной, на 0,2; 0,3; 0,6 и 0,5 %, соответственно. При этом хотелось бы отметить, что корковая часть надпочечников принимает важное участие в приспособительных реакциях организма к факторам окружающей среды, а их гормоны (адреналин и норадреналин) оказывают влияние на обмен веществ в тканях печени и мышц, а также поддерживают водный обмен и кровообращение
в почках. Масса фабрициевой сумки, являющаяся местом активного образования антителопродуцентов, в опытных группах незначительно превосходила изучаемый показатель в контрольной группе на 3,4; 5,2; 7,5 и 7,9 %. Поджелудочная железа, обладающая внешней и внутренней секрецией (вырабатывает панкреатический сок, участвующий в расщеплении белков, жиров, углеводов,
а также за счет выработки гормонов (инсулин и глюкоген) регулирует углеводный обмен), в опытных группах была по массе больше, чем в контрольной на 1,4; 3,5; 4,9 и 4,2 %, соответственно.
Полученные данные свидетельствуют о том, что цыплята-бройлеры, в рационе которых применяли пробиотик Галлобакт-Ф-Плюс, были меньше подвержены влиянию неблагоприятных факторов окружающей среды, а также
у них лучше протекали процессы обмена веществ, усиление роста и развития птицы, повышалась мясная продуктивность.
Анализ химического состава мяса цыплят представлен в таблице 83.
Таблица 83 – Химический состав мяса цыплят-бройлеров кросса Росс 308
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Грудные мышцы | |||||
| Влага, % | 70,94±1,13 | 70,94±1,21 | 70,90±1,20 | 70,84±1,31 | 70,90±1,24 |
| Белок, % | 23,24±0,53 | 23,28±0,47 | 23,36±0,51 | 23,52±0,56 | 23,40±0,50 |
| Жир, % | 4,65±0,16 | 4,60±0,13 | 4,55±0,15 | 4,46±0,10 | 4,51±0,18 |
| Зола, % | 1,17±0,03 | 1,18±0,02 | 1,19±0,02 | 1,18±0,03 | 1,19±0,03 |
| Индекс качества мяса |
0,20±0,01 | 0,20±0,01 | 0,19±0,01 | 0,18±0,01 | 0,19±0,01 |
| Ножные мышцы | |||||
| Влага, % | 71,84±1,36 | 71,78±1,31 | 71,76±1,37 | 71,72±1,28 | 71,72±1,33 |
| Белок, % | 22,32±0,51 | 22,39±0,49 | 22,47±0,58 | 22,57±0,47 | 22,53±0,52 |
| Жир, % | 4,78±0,13 | 4,75±0,15 | 4,70±0,13 | 4,64±0,11 | 4,67±0,16 |
| Зола, % | 1,06±0,02 | 1,08±0,03 | 1,07±0,03 | 1,07±0,02 | 1,08±0,03 |
| Индекс качества мяса |
0,21±0,01 | 0,21±0,01 | 0,20±0,01 | 0,20±0,01 | 0,20±0,01 |
Анализ химического состава грудных мышц цыплят-бройлеров не показал статистически достоверных различий в разрезе контрольной и опытных групп, однако по отдельным показателям была выявлена положительная тенденция от применения биопрепарата.
Так, содержание белка в грудных мышцах птиц 1–4-й опытных групп было выше, чем в контрольной, на 0,04; 0,12; 0,26 и 0,16 %, а уровень жира ниже на 0,05; 0,10; 0,19 и 0,14 %. Индекс качества мяса в опытных группах превосходил изучаемый показатель в контрольной группе. При этом во 2-й опытной группе он составил 0,19 ед., в 3-й опытной группе – 0,18 ед., в 4-й опытной группе – 0,19 ед., а в контрольной – 0,20 ед. Химический состав ножных мышц опытных групп также обладал наилучшими показателями по сравнению с контрольной группой. Уровень белка в 1-й опытной группе составил 22,39 %, во 2-й группе – 22,47 %, в 3-й опытной – 22,57 % и в 4-й опытной – 22,53%, что на 0,07; 0,15; 0,25 и 0,23 % выше, чем в контрольной группе. Содержание жира было в опытных группах ниже на 0,03; 0,08; 0,14 и 0,11 %, соответственно. Индекс качества мяса во 2-й, 3-й и 4-й опытных группах был ниже, чем в контрольной, и составил 0,20 ед., против 0,21 ед. в контроле. Полученные результаты говорят о том, что качество мяса цыплят-бройлеров опытных групп было не только более биополноценным за счет высокого содержания белка, но и обладает наилучшими диетическими свойствами за счет низкого уровня жира в мышечной ткани.
Полноценность мясной продукции цыплят определяли по показателю аминокислотного состава мышц птиц (рисунок 33).
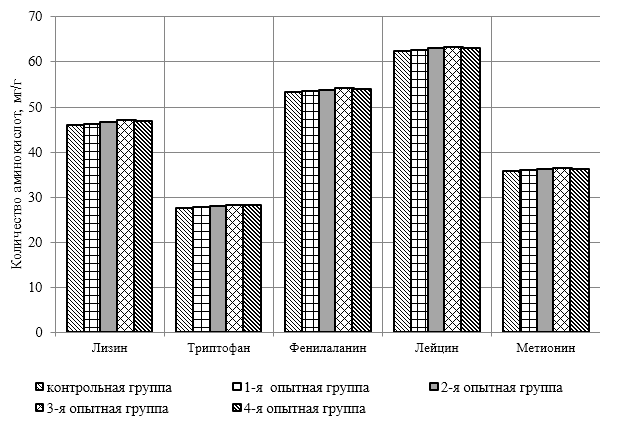
Рисунок 33 – Содержание отдельных незаменимых аминокислот
в мышцах птиц, мг/г
Положительная динамика от применения пробиотика была обнаружена при анализе аминокислотного скора мясной продукции цыплят-бройлеров. Установлено, что содержание лизина было выше в опытных группах по сравнению с контрольной на 0,9; 1,6; 2,9 и 2,2 %; триптофана – на 1,0; 2,1; 2,8 и 2,4 %; фенилаланина – на 0,3; 1,0; 1,7 и 1,3 %; лейцина – на 0,5; 1,0; 1,5 и 1,2 %; метионина – на 0,3; 1,1; 2,0 и 1,3 %, соответственно.
Изучение вкусовых качеств продукции бройлерного производства осуществлялась дегустационной оценкой варенного мяса подопытных птиц, а также бульона из него (таблица 84).
Таблица 84 – Дегустационная оценка мяса цыплят-бройлеров и бульона, баллы
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Грудные мышцы | 4,8±0,1 | 4,8±0,2 | 4,8±0,1 | 4,9±0,1 | 4,9±0,2 |
| Ножные мышцы | 4,9±0,2 | 4,9±0,1 | 4,9±0,2 | 4,9±0,2 | 4,9±0,1 |
| Бульон | 4,7±0,1 | 4,7±0,2 | 4,8±0,1 | 4,9±0,1 | 4,9±0,1 |
Бульон, полученный при варке мяса исследуемых групп был наваристым, соломенного цвета, обладал приятным вкусом и ароматом, характерным для мяса с.-х. птицы. На поверхности бульона обнаруживались капли жира. Варенное мясо было приятным, ароматным, нежным и сочным. Посторонние запахи ‒ отсутствовали. По совокупности показателей дегустационная комиссия отдала предпочтение мясу и бульону от птиц, полученных из 3-й и 4-й опытных групп.
В качестве завершающей оценки качества мясной продукции птицеводства была проведена ветеринарно-санитарная экспертиза, которая подтвердила её безопасность для использования в пищу (таблица 85).
Таблица 85 – Физико-химические и микробиологические показатели качества мяса цыплят-бройлеров
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Р-ция с сернокислой медью | – | – | – | – | – |
| Р-ция с формалином | – | – | – | – | – |
| Р-ция на пероксидазу | + | + | + | + | + |
| Количество ЛЖК, мг КОН/100 г | 1,54±0,03 | 1,51±0,03 | 1,50±0,02 | 1,45±0,02 | 1,47±0,02 |
| Количество микробных клеток в одном поле зрения микроскопа: |
|||||
| – с поверхности тушки | 3,56±0,09 | 3,48±0,11 | 3,42±0,10 | 3,35±0,10 | 3,38±0,09 |
| – с глубоких слоев | – | – | – | – | – |
| рН мяса, ед. | |||||
| Первые сутки | 6,87±0,20 | 6,86±0,16 | 6,84±0,15 | 6,89±0,19 | 6,88±0,19 |
| Вторые сутки | 6,45±0,21 | 6,40±0,22 | 6,44±0,22 | 6,42±0,24 | 6,41±0,22 |
| Третьи сутки | 6,08±0,17 | 6,05±0,16 | 6,02±0,18 | 6,01±0,18 | 6,01±0,17 |
Из таблицы 85 видно, что мясо птицы, полученное от изучаемых групп, было свежим и без признаков патологии, так как реакция с сернокислой медью и формалином была отрицательной, а на пероксидазу – положительной. Количество летучих жирных кислот и микробных клеток не выходило за пределы требований нормативной документации. Значение кислотности мяса в течение трех суток во всех группах снижалось до показателей, характерных для свежего и здорового мяса.
Таким образом, применение в составе рациона цыплят-бройлеров кросса Росс 308 пробиотика Галлобакт-Ф-Плюс способствует улучшению качественных показателей мясной продукции птицеводства, а также повышению сохранности, росту птицы и иммунологического статуса организма. Наилучшие результаты получены в 3-й опытной группе с дозировкой пробиотика равной 0,75 мл/гол.
Экономическая эффективность использования пробиотика. Результаты экономической эффективности использования пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах кросса Росс 308 представлены в таблице 86.
В результате проведенного предварительного анализа экономической эффективности применения в рационе цыплят-бройлеров кросса Росс 308 пробиотической добавки Галлобакт-Ф-Плюс выявлено, что прибыль в 1–4-й опытных группах составила 3 201,96; 3 290,45; 3 615,84 и 3 252,26 руб., против 2 898,29 руб. в контрольной группе, что выше на 303,67; 392,16; 717,55
и 353,97 руб., соответственно. В целом, наилучшие результаты были получены в 3-й опытной группе, в которой экономический эффект превышал контрольную группу на 24,76 %.
Таким образом, экономически целесообразно проводить производственную апробацию пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах кросса Росс 308 в дозе, составляющей 0,75 мл/гол.
Влияние пробиотика Галлобакт-Ф-Плюс на цыплят-бройлеров
кросса Кобб 500
Для изучения влияния пробиотика Галлобакт-Ф-Плюс на сохранность, продуктивность, основные показатели обмена веществ, а также качество продукции от цыплят-бройлеров кросса Кобб 500 был проведен научно-хозяйственный опыт, в котором методом групп-аналогов было сформировано пять групп птиц по 60 гол. в каждой: контрольная группа – птица получала стандартный рацион, рекомендованный ВНИТИП, и имела свободный доступ к воде; первая опытная группа – подопытная птица получала стандартный рацион,
а также дополнительно в воду вводился пробиотик Галлобакт-Ф-Плюс в дозе 0,25 мл/гол.; вторая опытная группа – Галлобакт-Ф-Плюс вводили в дозе 0,5 мл/гол.; третья опытная группа – объекты исследований получали стандартный рацион, рекомендованный ВНИТИП, а также в воду добавляли пробиотик в дозе 0,75 мл/гол.; четвертая опытная группа – объекты исследований получали стандартный рацион, а в воду добавляли пробиотик в дозе 1,0 мл/гол. Опыт продолжался 42 дня.
Рост и развитие цыплят-бройлеров. Результаты хозяйственных показателей цыплят-бройлеров представлены в таблице 87 и на рисунке 34.
Таблица 87 – Изучаемые хозяйственные показатели цыплят-бройлеров
кросса Кобб 500 (n = 60)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Сохранность, % | 95,0 | 98,3 | 100 | 100 | 100 |
| Прирост живой массы за период выращивания (042 сут) | |||||
| Одной головы в среднем, г |
2572,10 | 2604,24 | 2622,12 | 2653,47 | 2645,50 |
| Расход комбикорма за период выращивания (042 сут) | |||||
| 1-й головы, г | 4587,81 | 4608,48 | 4625,78 | 4628,59 | 4627,93 |
| Конверсия, кг | 1,78 | 1,77 | 1,76 | 1,74 | 1,75 |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
При изучении хозяйственных показателей цыплят-бройлеров кросса Кобб 500 были получены аналогичные результаты как на кроссе Росс 308. Применение пробиотика в зависимости от дозы способствовало равномерному превосходству живой массы птицы в опытных группах по сравнению с контрольной. Незначительные изменения по живой массе цыплят-бройлеров были зафиксированы уже на 7-е сутки ее взвешивания, где в 1–4-й опытных группах изучаемый показатель превысила контрольную группу на 0,7; 0,9; 2,0 и 1,3 %, соответственно, но разница была не достоверна.
Таблица 86 – Экономическая эффективность использования пробиотика Галлобакт-Ф-Плюс
| Показатель | Группа | |||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Поголовье в начале опыта, гол. | 60 | 60 | 60 | 60 | 60 | |
| Сохранность, % | 93,3 | 96,6 | 96,6 | 100 | 100 | |
| Поголовье в конце опыта, гол. | 56 | 58 | 58 | 60 | 60 | |
| Масса тушки на реализацию | ||||||
| Масса потрошенной тушки, г | 1813,27±10,56 | 1853,23±10,87 | 1873,76±11,04 | 1912,42±11,35* | 1903,82±11,42* | |
| Масса потрошенной тушки всего, кг | 101,54 | 107,49 | 108,68 | 114,75 | 114,23 | |
| Затраты комбикорма за период выращивания | ||||||
| На голову, г | 4798,74 | 4814,25 | 4828,76 | 4836,41 | 4843,68 | |
| На все поголовье, кг | 268,73 | 279,23 | 280,07 | 290,18 | 290,62 | |
| Затраты пробиотика | ||||||
| Всего, мл | – | 130,5 | 261,0 | 405,0 | 540,0 | |
| Экономическая эффективность | ||||||
| Цена 1 кг комбикорма, руб. | 27 | |||||
| Стоимость израсходованного корма всего, руб. |
7 255,71 | 7 539,21 | 7 561,89 | 7 834,86 | 7 846,74 | |
| Цена 1 л пробиотика, руб. | – | 60 | ||||
| Стоимость пробиотика всего, руб. | – | 7,83 | 15,66 | 24,30 | 324,00 | |
| Стоимость израсходованного корма и пробиотика, руб. |
7 255,71 | 7 547,04 | 7 577,55 | 7 859,16 | 8 170,74 | |
| Цена реализации 1 кг мяса птицы, руб. | 100,0 | |||||
| Выручка от реализации мяса птицы, руб. | 10 154,00 | 10 749,00 | 10 868,00 | 11 475,00 | 11 423,00 | |
| Прибыль от реализации мяса птицы, руб. | 2 898,29 | 3 201,96 | 3 290,45 | 3 615,84 | 3 252,26 | |
| Экономический эффект от использования пробиотика к контролю,
руб. % |
–
– |
303,67
10,48 |
392,16
13,53 |
717,55
24,76 |
353,97
12,21 |
|
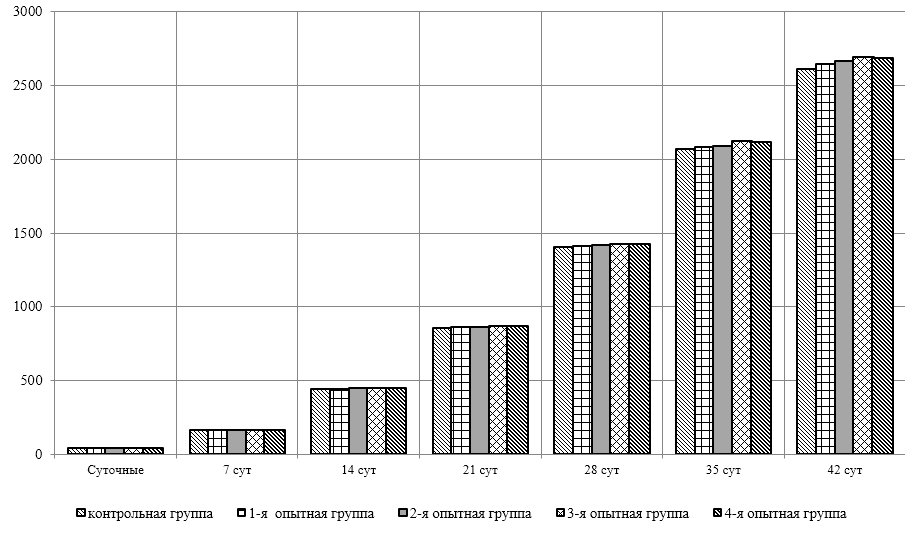
Рисунок 34 ‒ Динамика живой массы, г
Статистически достоверная разница по массе цыплят-бройлеров была выявлена на 35-й день взвешивания в 3-й и 4-й опытных группах по сравнению с контрольной, в которых данный показатель был выше на 57,40
и 49,14 г, или 2,8 и 2,4 % (Р < 0,05). На 42-е сутки взвешивания птицы живая масса во 2–4-й опытных группах превысила изучаемый показатель в контрольной на 1,9; 3,1 и 2,8 % при достоверной разнице (Р < 0,05).
Жизнеспособность птиц оценивали по показателю сохранности, который
в контрольной группе составил 95,0 %, в то время как в 1-й опытной группе он был 98,3 %, а во 2-й, 3-й и 4-й опытных группах – 100,0 %.
Анализируя прирост массы цыплят-бройлеров за весь период исследований установлено, что данный показатель в опытных группах превысил контроль на 32,14 г (1,2 %), 50,02 г (1,9 %), 81,37 г (3,1 %) и 73,40 г (2,9 %), соответственно.
Показатель конверсии корма в опытных группах был ниже, чем в контрольной и составил в 1-й опытной – 1,77 кг, во 2-й опытной – 1,76 кг, в 3-й
и 4-й опытных – 1,74 и 1,75 кг, соответственно. Против 1,78 кг в контрольной группе.
Таким образом, применение пробиотика Галлобакт-Ф-Плюс положительно сказалось на хозяйственных показателях цыплят-бройлеров кросса Кобб 500, особенно при дозе биопрепарата 0,75 мл/гол.
Морфологические и биохимические показатели крови. Результаты морфологических и биохимических показателей крови цыплят-бройлеров кросса Кобб 500 представлены в таблице 88.
Из таблицы 88 видно, что применение пробиотика не имело существенного влияния на морфо-биохимический статус крови цыплят-бройлеров кросса Кобб 500 на 21-е сутки исследования. Все показатели в подопытных группах находились на уровне физиологической нормы. Иная картина была установлена при анализе крови на 42-е сутки, где была зафиксирована положительная тенденция по изучаемым показателям в опытных группах. Так, количество эритроцитов во 2-й, 3-й и 4-й опытных группах превысило аналогичный показатель в контрольной на 4,5 и 8,2 % при статистически достоверной разнице (Р < 0,05), при этом в 1-й опытной группе также было превосходство на 1,5 %, но без достоверной разницы.
Таблица 88 – Морфо-биохимический статус крови цыплят-бройлеров
кросса Кобб 500
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| 21-е сутки | |||||
| Эритроциты, 1012/л | 2,58±0,03 | 2,61±0,04 | 2,62±0,04 | 2,66±0,04 | 2,65±0,03 |
| Гемоглобин, г/л | 84,23±1,41 | 84,78±1,50 | 84,68±1,54 | 85,05±1,49 | 84,86±1,55 |
| Тромбоциты, 109/л | 63,56±0,81 | 63,78±0,78 | 63,95±0,77 | 64,08±0,74 | 63,94±0,80 |
| Лейкоциты, 109/л | 24,65±0,43 | 24,49±0,41 | 24,54±0,36 | 24,58±0,45 | 24,61±0,38 |
| Общий белок, г/л | 37,56±0,51 | 37,71±0,57 | 38,32±0,53 | 38,45±0,49 | 38,38±0,46 |
|
15,25±0,21 | 15,32±0,17 | 15,36±0,19 | 15,38±0,17 | 15,35±0,21 |
|
22,31±0,32 | 22,39±0,29 | 22,96±0,35 | 23,07±0,31 | 23,03±0,28 |
| А/Г коэффициент | 0,68±0,02 | 0,68±0,03 | 0,67±0,02 | 0,67±0,01 | 0,67±0,02 |
| Холестерин, мМ/л | 3,43±0,04 | 3,47±0,03 | 3,41±0,04 | 3,40±0,05 | 3,41±0,03 |
| АСТ, Ед/л | 205,58±2,63 | 207,48±2,53 | 206,38±2,62 | 205,39±2,57 | 206,87±2,55 |
| АЛТ, Ед/л | 23,78±0,29 | 23,58±0,24 | 23,52±0,25 | 23,61±0,23 | 23,61±0,27 |
| Кальций, мМ/л | 9,41±0,12 | 9,43±0,12 | 9,42±0,10 | 9,48±0,09 | 9,46±0,11 |
| Фосфор, мМ/л | 4,43±0,06 | 4,47±0,07 | 4,48±0,05 | 4,49±0,08 | 4,45±0,09 |
| БАСК, % | 38,7±1,74 | 38,8±1,81 | 39,3±1,77 | 39,5±1,69 | 39,2±1,72 |
| ЛАСК, % | 33,5 ±1,31 | 33,6±1,21 | 33,8±1,25 | 34,1±1,28 | 33,8±1,24 |
| 42-е сутки | |||||
| Эритроциты, 1012/л | 2,69±0,01 | 2,73±0,03 | 2,81±0,02* | 2,91±0,03* | 2,91±0,02* |
| Гемоглобин, г/л | 85,41±0,81 | 86,13±0,79 | 87,54±0,93 | 89,37±0,72* | 89,13±0,77* |
| Тромбоциты, 109/л | 65,02±0,51 | 66,41±0,60 | 67,44±0,54 | 68,73±0,55* | 68,68±0,50* |
| Лейкоциты, 109/л | 25,70±0,42 | 25,16±0,39 | 26,02±0,38 | 26,21±0,40 | 26,08±0,38 |
| Общий белок, г/л | 38,31±0,28 | 39,02±0,31 | 39,57±0,27 | 40,71±0,23* | 40,39±0,23* |
|
15,21±0,20 | 15,54±0,21 | 16,11±0,20 | 17,27±0,20* | 17,22±0,19* |
|
23,10±0,32 | 23,48±0,31 | 23,46±0,27 | 23,44±0,29 | 23,17±0,29 |
| А/Г коэффициент | 0,65±0,02 | 0,66±0,01 | 0,68±0,03 | 0,73±0,03 | 0,74±0,03 |
| Холестерин, мМ/л | 3,69±0,02 | 3,65±0,03 | 3,58±0,04 | 3,51±0,02* | 3,52±0,01* |
| АСТ, Ед/л | 225,31±2,73 | 224,74±2,69 | 225,11±2,63 | 223,74±2,71 | 223,92±2,70 |
| АЛТ, Ед/л | 24,18±0,32 | 23,89±0,41 | 24,03±0,35 | 29,91±0,27 | 24,01±0,33 |
| Кальций, мМ/л | 9,37±0,10 | 9,41±0,11 | 9,57±0,09 | 9,63±0,08 | 9,61±0,10 |
| Фосфор, мМ/л | 4,24±0,04 | 4,28±0,05 | 4,33±0,04 | 4,39±0,03 | 4,40±0,06 |
| БАСК, % | 42,04±0,31 | 43,02±0,27 | 43,71±0,28 | 44,87±0,31* | 44,72±0,25* |
| ЛАСК, % | 33,11±0,21 | 33,73±0,19 | 34,22±0,24 | 35,72±0,22* | 35,51±0,21* |
| Примечание: * – разница с контролем достоверна (Р < 0,05). | |||||
Статистически достоверная разница проявилась при анализе гемоглобина, уровень которого в 3-й и 4-й опытных группах был выше, чем в контрольной на 4,6 и 4,4 %; тромбоцитов больше на 5,7 и 5,6%; общего белка – на 6,3 и 5,4 %; альбуминов – на 13,5 и 13,2 %; холестерина ниже на 4,9 и 4,6 %; бактерицидная активность выше на 2,83 и 2,68%; лизоцимная активность – на 2,61 и 2,4 %, соответственно (Р < 0,05). Во всех опытных группах было выявлено превосходство по сравнению с контрольной группой в альбумино-глобулиновом коэффициенте, значение которого составило 0,66; 0,68; 0,73 и 0,74 ед., против 0,65 ед. в контроле. Была выявлена тенденция в превосходстве по отношению к контрольной группе содержания в сыворотки крови цыплят-бройлеров опытных групп кальция и фосфора, соответственно, на 0,4 и 0,9 % (1-я опытная группа); 2,1 и 2,1 % (2-я опытная группа); 2,8 и 3,5 % (3-я опытная группа); 2,6 и 3,8 % (4-я опытная группа).
Таким образом, пробиотик Галлобакт-Ф-Плюс идентично цыплятам-бройлерам кросса Росс 308 оказывает на кросс Кобб 500 положительное влияние через стимуляцию обменных процессов, с одновременным усилением иммунного потенциала птицы, что и обеспечивает их высокую сохранность
и продуктивность.
Влияние пробиотика на пищеварение. Данное влияние оценивали по показателю переваримости компонентов кормосмеси, для чего проводили балансовый опыт в конце выращивания птицы, результаты которого продемонстрированы на рисунке 35.
Из данных рисунка 35 установлено, что применение в составе рациона птицы опытных групп пробиотика оказала наибольший эффект по сравнению с контрольной группой, что выразилось в статистически достоверном повышении
в 3-й и 4-й опытных группах переваримости сырого протеина на 5,08 и 4,76 %,
а также сырой клетчатки – на 2,37 и 2,04 % (Р < 0,05). Была выявлена положительная тенденция в переваримости сырого жира, которая превысила в опытных группах данный показатель контрольной, на 0,42; 1,14; 2,34 и 1,87 %, соответственно. Показатель переваримости БЭВ в опытных группах также был выше, чем в контрольной, и составил в 1-й опытной группе – 52,16 %, во 2-й опытной 53,02 %, в 3-й опытной 54,38 % и в 4-й опытной 54,24%, против 51,37 % в группе контроля.
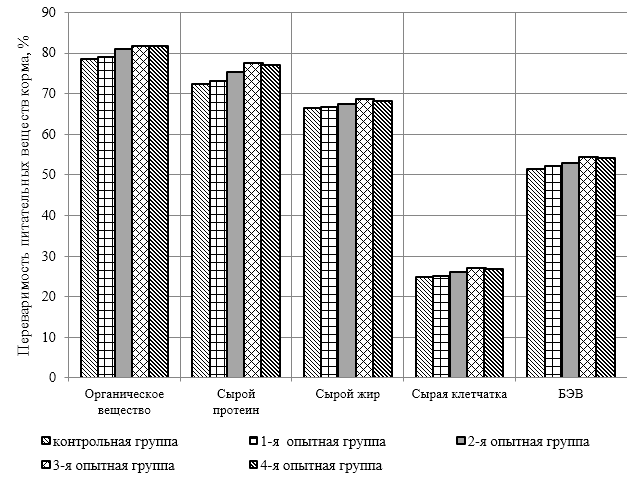
Рисунок 35 – Переваримость питательных веществ комбикорма
цыплятами-бройлерами кросса Кобб 500, %
Таким образом, применение в составе рациона цыплят-бройлеров кросса Кобб 500 пробиотика Галлобакт-Ф-Плюс поспособствовало лучшему перевариванию веществ комбикорма, что обеспечило повышение продуктивности птицы опытных групп.
Микробиоценноз химуса кишечника. Результаты микробиологических исследований слепых отростков кишечника цыплят-бройлеров кросса Кобб 500 после использования в их рационе пробиотика Галлобакт-Ф-Плюс представлены в таблице 89.
Использование пробиотика положительно сказалось на микробном балансе кишечника цыплят-бройлеров всех опытных групп. Так, уровень молочнокислых микроорганизмов и бифидобактерий был статистически достоверно выше
в 1-й опытной группе по сравнению с контрольной в 1,1 и 1,0 раза; во 2-й опытной группе – в 1,3 и 1,1 раза; в 3-й опытной – 1,5 и 1,3 раза; в 4-й опытной – в 1,4 и 1,2 раза (P < 0,05). Также в опытных группах достоверно снижался титр эшерихий, энтерококков, стафилококков и стрептококков. В изучаемых опытных группах и контрольной не было зарегистрировано в химусе псевдомонад и клостридий. В 3-й и 4-й опытных группах отсутствовали представители энтерококков.
Таблица 89 – Баланс полезной и условно-патогенной микрофлоры химуса
слепых отростков цыплят-бройлеров кросса Кобб 500 (n = 10)
| Показатели | Группы | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Молочнокислые бактерии, 108 КОЕ/г |
2,80,04 | 3,10,03* | 3,70,05* | 4,20,04* | 4,10,06* |
| Бифидобактерии, 108 КОЕ/г | 2,90,02 | 3,00,03* | 3,20,04* | 3,80,03* | 3,70,04* |
| Эшерихии, 103 КОЕ/г | 5,30,06 | 5,00,05* | 4,30,05* | 3,30,03* | 3,50,04* |
| Энтерококки, 102 КОЕ/г | 4,90,05 | 3,30,03* | 4,80,02* | ‒ | ‒ |
| Псевдомонады, 10 КОЕ/г | ‒ | ‒ | ‒ | ‒ | ‒ |
| Стафилококки, 102 КОЕ/г | 5,70,05 | 5,10,04* | 4,70,03* | 3,80,04* | 3,80,04* |
| Стрептококки, 102 КОЕ/г | 6,10,06 | 5,60,04* | 5,10,04* | 4,50,03* | 4,60,05* |
| Клостридии, 10 КОЕ/г | ‒ | ‒ | ‒ | ‒ | ‒ |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Мясная продуктивность, развитие внутренних органов и качество мяса птиц. Результаты убойного выхода цыплят-бройлеров кросса Кобб 500 представлены в таблице 90.
Из таблицы 90 видно, что за счет статистически достоверного повышения живой массы тела цыплят-бройлеров во 2–4-й опытных группах наблюдалось достоверная разница по массе потрошенной тушки, которая превысила данное значение в контрольной группе на 2,7; 4,8 и 3,9 % (P < 0,05). Убойный выход
в опытных группах был выше, чем в контрольной, на 0,2; 0,7; 1,4 и 1,0 %, соответственно.
Проводилось изучение влияния пробиотика на соотношение мышечной, кожной и костной тканей составных частей тела тушки (грудь, бедро, голень). Результаты разделки грудной части представлены в таблице 91.
Таблица 90 – Убойные показатели цыплят-бройлеров кросса Кобб 500 (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | |
| Предубойная масса птицы, г | 2620,727,12 | 2649,116,73 | 2665,516,81* | 2693,326,80* | 2685,286,83* |
| Масса потрошеной тушки, г | 1810,927,54 | 1835,837,31 | 1860,537,45* | 1898,797,42* | 1882,387,37* |
| Убойный выход, % | 69,1 | 69,3 | 69,8 | 70,5 | 70,1 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | |||||
Таблица 91 – Морфологический состав грудки цыплят-бройлеров (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Масса потрошеной тушки | г | 1810,927,54 | 1835,837,31 | 1860,537,45* | 1898,797,42* | 1882,387,37* |
| Масса грудных мышц | г | 398,403,44 | 405,713,72 | 416,764,02 | 431,034,07* | 425,403,77* |
| % | 22,0 | 22,1 | 22,4 | 22,7 | 22,6 | |
| Масса кожи | г | 56,1410,92 | 55,070,94 | 59,540,95 | 58,860,98 | 58,351,10 |
| % | 3,1 | 3,0 | 3,2 | 3,1 | 3,1 | |
| Масса костей | г | 68,810,9 | 71,601,01 | 72,560,98 | 74,051,09 | 73,411,07 |
| % | 3,8 | 3,9 | 3,9 | 3,9 | 3,9 | |
| Масса всех тканей | г | 523,355,63 | 533,185,21 | 548,865,42 | 563,945,70* | 557,165,54* |
| % | 28,9 | 29,0 | 29,5 | 29,7 | 29,6 | |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||||||
Масса грудных мышц в 1-й и 2-й опытных группах имела незначительную тенденцию к увеличению по сравнению с контрольной на 7,31 и 18,36 г. В 3-й
и 4-й опытных по данному показателю наблюдалась статистически достоверное превосходство по сравнению с контрольной группой на 8,2 и 6,8 % (P < 0,05). На долю грудных мышц по отношению к потрошённой тушке в контрольной группе приходится 22,0 %, в то время как в 1-й опытной – 22,1 %, во 2-й опытной – 22,4 %, в 3-й опытной –22,7%, а в 4-й опытной – 22,6 %. Масса кожи грудки 2–4-й опытных групп незначительно превышала данный показатель
в контрольной группе, но без достоверной разницы. В целом, мышечная, кожная и костная ткани опытных групп были больше по сравнению с контрольной группой, но статистически достоверные различия наблюдались в 3-й и 4-й опытных группах, разница составила 40,62 и 33,84 г, или 7,8 и 6,5 % (P < 0,05).
Результаты морфологического состава бедра цыплят-бройлеров кросса Кобб 500 представлены в таблице 92.
Изучение бедренной части тушки цыплят-бройлеров кросса Кобб 500 показали, что масса бедренных мышц в 1–4-й опытных группах составила 223,97; 228,85; 235,45 и 231,53 г, что на 1,4; 3,6; 6,6 и 4,8 % выше, чем в контрольной группе. Выход мяса в разрезе групп был почти равнозначным и составил 12,2; 12,3; 12,4 и 12,3 % в опытных группах, против 12,2 % в контрольной. По массе всех тканей бедренной части установлено, что только в 3-й опытной группе было зафиксировано статистически достоверное повышение показателя по сравнению с контрольной группой на 7,4 % (P < 0,05), в остальных опытных группах наблюдалась тенденция к повышению изучаемого показателя.
Морфологический состав голени цыплят-бройлеров кросса Кобб 500 представлен в таблице 93.
Масса мышц голени в опытных группах, аналогично бедренным, превышала полученные значения в контрольной группе на 1,4; 4,0; 6,1 и 5,2%, соответственно. По сумме всех составляющих частей голени наибольшее значение было выявлено в 3-й опытной группе, которое составило 267,72 г, затем в 4-й опытной – 265,42 г, во 2-й опытной – 262,34 г и в 1-й опытной – 255,18 г против 251,72 г в контрольной группе.
Таблица 92 – Морфологический состав бедра цыплят-бройлеров (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Масса потрошеной тушки | г | 1810,92±7,54 | 1835,83±7,31 | 1860,53±7,45* | 1898,79±7,42* | 1882,38±7,37 |
| Масса бедренных мышц | г | 220,93±2,51 | 223,97±2,72 | 228,85±2,67 | 235,45±2,69 | 231,53±2,62 |
| % | 12,2 | 12,2 | 12,3 | 12,4 | 12,3 | |
| Масса кожи | г | 38,03±0,61 | 40,39±0,63 | 40,93±0,65 | 43,67±0,66* | 43,29±0,65* |
| % | 2,1 | 2,2 | 2,2 | 2,3 | 2,3 | |
| Масса костей | г | 43,46±0,61 | 44,49±0,51 | 44,65±0,57 | 45,57±0,60 | 45,17±0,58 |
| % | 2,4 | 2,4 | 2,4 | 2,4 | 2,4 | |
| Масса всех тканей | г | 302,43±3,43 | 308,85±3,52 | 314,43±3,44 | 324,69±3,43* | 319,99±3,48 |
| % | 16,7 | 16,8 | 16,9 | 17,1 | 17,0 | |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||||||
Таблица 93 – Морфологический состав голени цыплят-бройлеров (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Масса потрошеной тушки | г | 1810,92±7,54 | 1835,83±7,31 | 1860,53±7,45* | 1898,79±7,42* | 1882,38±7,37 |
| Масса мышц голени | г | 155,74±2,77 | 157,88±2,71 | 161,87±2,82 | 165,19±2,80 | 163,77±2,79 |
| % | 8,6 | 8,6 | 8,7 | 8,7 | 8,7 | |
| Масса кожи | г | 38,03±0,62 | 38,55±0,69 | 40,93±0,72 | 41,77±0,66 | 41,41±0,65 |
| % | 2,1 | 2,1 | 2,2 | 2,2 | 2,2 | |
| Масса костей | г | 57,95±0,74 | 58,75±0,71 | 59,54±0,77 | 60,76±0,78 | 60,24±0,76 |
| % | 3,2 | 3,2 | 3,2 | 3,2 | 3,2 | |
| Масса всех тканей | г | 251,72±3,31 | 255,18±3,21 | 262,34±3,34 | 267,72±3,37 | 265,42±3,29 |
| % | 13,9 | 13,9 | 14,1 | 14,1 | 14,1 | |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||||||
Результаты развития внутренних органов цыплят-бройлеров кросса Кобб 500 представлены в таблице 94.
Несмотря на то, что соотношение органов в разрезе изучаемых подопытных групп к массе потрошенной тушки были почти идентичны, их масса имела незначительную разницу. Например, масса печени в 1–4-й опытных группах была больше, чем в контрольной группе на 0,14; 0,34; 0,85 и 0,67 г; сердце – на 0,33; 0,62; 0,86 и 0,85 г, соответственно. Данные различия не имели достоверной разницы, патологии не выявлено.
Результаты влияния пробиотика Галлобакт-Ф-Плюс на развитие желез внутренней секреции у цыплят-бройлеров кросса Кобб 500 представлены в таблице 95.
Таблица 95 – Развитие эндокринных желез цыплят-бройлеров, мг (n = 10)
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Щитовидная железа |
181,18±1,83 | 181,27±1,74 | 181,54±1,81 | 181,71±1,77 | 181,72±1,80 |
| Паращитовидная железа | 14,52±0,63 | 14,74±0,61 | 14,84±0,57 | 14,91±0,60 | 14,87±0,56 |
| Надпочечник | 163,74±2,11 | 163,92,=2,02 | 164,12±1,98 | 164,20±1,99 | 164,02±2,04 |
| Фабрициева сумка |
2,61±0,05 | 2,67±0,03 | 2,72±0,05 | 2,78±0,06 | 2,77±0,08 |
| Поджелудочная железа | 2,73±0,05 | 2,79±0,04 | 2,82±0,06 | 2,91±0,03 | 2,88±0,06 |
Использование в рационе цыплят-бройлеров опытных групп пробиотика Галлобакт-Ф-Плюс не имело достоверного влияния на массу эндокринных желез, однако была выявлена незначительная тенденция к повышению изучаемых показателей. Так, масса щитовидной железы, паращитовидной, надпочечника, фабрициевой сумки и поджелудочной железы в 1-й опытной группе были выше, чем в контрольной, на 0,04; 1,5; 0,1; 2,3 и 2,2 %; во 2-й опытной – на 0,2; 2,2; 0,2; 4,2 и 3,3 %; в 3-й опытной – на 0,3; 2,7; 0,3; 6,5 и 3,3 %; в 4-й опытной – на 0,3; 2,4; 0,2; 6,1 и 5,5 %.
Таблица 94 – Развитие внутренних органов цыплят-бройлеров (n = 10)
| Показатель | Ед. изм. | Группа | ||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Предубойная масса птицы | г | 2620,72±7,12 | 2649,11±6,73 | 2665,51±6,81* | 2693,32±6,80* | 2685,28±6,83* |
| Печень | г | 35,17±0,54 | 35,31±0,61 | 35,51±0,59 | 36,02±0,67 | 35,84±0,64 |
| % | 1,34 | 1,33 | 1,33 | 1,34 | 1,33 | |
| Сердце | г | 11,17±0,29 | 11,50±0,31 | 11,79±0,37 | 12,03±0,34 | 12,02±0,33 |
| % | 0,43 | 0,43 | 0,44 | 0,44 | 0,44 | |
| Мышечный желудок | г | 30,57±0,70 | 30,89±0,67 | 31,42±0,66 | 31,94±0,73 | 31,84±0,71 |
| % | 1,17 | 1,17 | 1,18 | 1,18 | 1,18 | |
| Железистый желудок | г | 10,07±0,28 | 10,22±0,32 | 10,41±0,37 | 10,94±0,29 | 10,83±0,30 |
| % | 0,38 | 0,38 | 0,40 | 0,40 | 0,40 | |
| Селезенка | г | 4,11±0,08 | 4,09±0,10 | 4,17±0,08 | 4,15±0,07 | 4,17±0,11 |
| % | 0,16 | 0,15 | 0,16 | 0,15 | 0,15 | |
| Кишечник | г | 116,56±1,42 | 117,13±1,34 | 117,81±1,49 | 118,76±1,42 | 118,61±1,44 |
| % | 4,44 | 4,42 | 4,42 | 4,41 | 4,42 | |
| Легкие | г | 10,35±0,40 | 10,54±0,35 | 10,84±0,39 | 11,10±0,33 | 11,05±0,38 |
| % | 0,39 | 0,40 | 0,40 | 0,41 | 0,41 | |
| Почки | г | 8,31±0,28 | 8,37±0,29 | 8,45±0,30 | 8,49±0,25 | 8,47±0,27 |
| % | 0,32 | 0,32 | 0,32 | 0,31 | 0,31 | |
| Желчный пузырь | г | 2,32±0,05 | 2,33±0,06 | 2,36±0,07 | 2,39±0,05 | 2,38±0,06 |
| % | 0,09 | 0,09 | 0,09 | 0,09 | 0,09 | |
Изучалось влияние пробиотика на химический состав мяса цыплят-бройлеров кросса Кобб 500, который представлен в таблице 96.
Таблица 96 – Химический состав мяса цыплят-бройлеров кросса Кобб 500
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Грудные мышцы | |||||
| Влага, % | 71,11±1,02 | 71,09±1,11 | 70,98±1,17 | 70,62±1,15 | 70,84±1,20 |
| Белок, % | 23,11±0,44 | 23,17±0,47 | 23,31±0,50 | 23,74±0,49 | 23,51±0,48 |
| Жир, % | 4,57±0,11 | 4,52±0,12 | 4,47±0,13 | 4,41±0,12 | 4,43±0,13 |
| Зола, % | 1,21±0,02 | 1,22±0,03 | 1,24±0,02 | 1,23±0,03 | 1,22±0,02 |
| Индекс качества мяса | 0,20±0,01 | 0,19±0,01 | 0,19±0,01 | 0,18±0,01 | 0,19±0,01 |
| Ножные мышцы | |||||
| Влага, % | 71,62±1,13 | 71,49±1,17 | 71,49±1,21 | 71,51±1,15 | 71,50±1,14 |
| Белок, % | 22,44±0,41 | 22,57±0,43 | 22,58±0,49 | 22,61±0,44 | 22,59±0,47 |
| Жир, % | 4,82±0,07 | 4,81±0,08 | 4,78±0,06 | 4,75±0,08 | 4,77±0,07 |
| Зола, % | 1,12±0,03 | 1,13±0,02 | 1,15±0,02 | 1,13±0,03 | 1,14±0,03 |
| Индекс качества мяса | 0,21±0,01 | 0,21±0,01 | 0,21±0,01 | 0,20±0,01 | 0,20±0,01 |
Результаты исследований в разрезе групп не показали статистически достоверных различий, но следует обратить внимание, что содержание белка
в грудных мышцах опытных групп было незначительно выше, чем в контрольной на 0,06; 0,2; 0,63 и 0,4 %, а в ножных – на 0,13; 0,14; 0,17 и 0,15 %, соответственно. Содержание жира в грудных мышцах опытных групп было незначительно ниже, чем в контрольной, на 0,05; 0,1; 0,16 и 0,14 %, а в ножных – на 0,01; 0,04; 0,07 и 0,05 %, соответственно. Индекс качества грудных мышц
в опытных группах составил 0,19 и 0,18 ед., против 0,20 ед. в контрольной группе. Индекс качества ножных мышц наилучшим был в 3-й и 4-й опытных группах и составил 0,20 ед., против 0,21 ед. в контрольной группе.
Биополноценность мышц цыплят-бройлеров кросса Кобб 500 определяли по аминокислотному скору (рисунок 36).
Результаты рисунка 36 показывают, что в белке мяса цыплят-бройлеров опытных групп содержалось более оптимальное количество незаменимых аминокислот. Так, уровень лизина в мышцах опытных групп был выше, чем контрольной на 0,1; 1,2; 2,5 и 2,0 %; триптофана – на 1,4; 3,6; 5,3 и 4,4 %; фенилаланина – на 0,1; 1,1; 2,4 и 1,9 %; лейцина – на 0,9; 1,8; 3,0 и 2,8 %; метионина – на 1,5; 3,3; 5,3 и 4,3 %, соответственно.
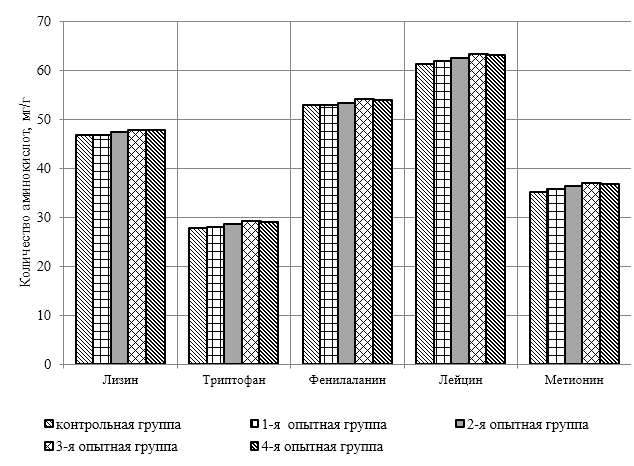
Рисунок 36 – Содержание отдельных незаменимых аминокислот
в мышцах птиц, мг/г
Органолептические качества мясной продукции определяли дегустационной оценкой, результаты которой представлены в таблице 97.
Результаты оценки в разрезе подопытных групп были высокими и существенной разницы не зафиксировано. Бульон, полученный при варке мяса контрольной и опытных групп, был наваристым, немного соломенного цвета, обладал приятным вкусом и ароматом, характерным для мяса птицы. Вареное мясо, полученное от контрольной и опытных групп, по вкусу было приятным, ароматным, нежным и умеренно сочным. Посторонние запахи – отсутствовали. Наивысшую оценку получили мясо и бульон во 2-й , 3-й и 4-й опытных группах.
Таблица 97 – Дегустационная оценка мяса цыплят-бройлеров и бульона, баллы
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Грудные мышцы | 4,8±0,1 | 4,8±0,1 | 4,9±0,1 | 4,9±0,2 | 4,9±0,1 |
| Ножные мышцы | 4,8±0,2 | 4,8±0,1 | 4,9±0,2 | 4,9±0,1 | 4,9±0,2 |
| Бульон | 4,8±0,2 | 4,8±0,2 | 4,9±0,2 | 4,9±0,1 | 4,9±0,1 |
Результаты ветеринарно-санитарной безопасности мясной продукции птицеводства представлены в таблице 98.
Таблица 98 – Физико-химические и микробиологические показатели качества мяса цыплят-бройлеров
| Показатель | Группа | ||||
| контрольная | 1-я опытная |
2-я опытная |
3-я опытная |
4-я опытная |
|
| Реакция с сернокислой медью | ‒ | ‒ | ‒ | ‒ | ‒ |
| Реакция с формалином | ‒ | ‒ | ‒ | ‒ | ‒ |
| Реакция на пероксидазу | + | + | + | + | + |
| Количество ЛЖК, мг КОН/100 г | 1,71±0,02 | 1,70±0,03 | 1,68±0,03 | 1,65±0,04 | 1,66±0,03 |
| Количество микробных клеток в одном поле зрения микроскопа:
|
3,42±0,07
‒ |
3,41±0,10
‒ |
3,38±0,08
‒ |
3,37±0,09
‒ |
3,38±0,09
‒ |
| рН мяса | |||||
| Первые сутки | 6,77±0,14 | 6,79±0,17 | 6,79±0,16 | 6,77±0,17 | 6,78±0,18 |
| Вторые сутки | 6,41±0,15 | 6,40±0,17 | 6,40±0,19 | 6,41±0,18 | 6,40±0,19 |
| Третьи сутки | 6,01±0,18 | 6,02±0,17 | 6,01±0,19 | 5,98±0,15 | 5,98±0,17 |
Как и в ранее проведенных экспериментах установлено, что по изучаемым физико-химическим и микробиологическим показателям мясо цыплят-бройлеров кросса Кобб 500 всех групп было безопасным для применения в пищу не зависимо от дозы пробиотика Галлобакт-Ф-Плюс.
Таким образом, по совокупности проведенных экспериментов выявлено, что пробиотик Галлобакт-Ф-Плюс оказывает положительное действие на организм цыплят-бройлеров кросса Кобб 500, стимулирует рост птицы, повышает сохранность, усиливает обмен веществ, повышает иммунный статус организма. Наилучшие показатели были выявлены при использовании биопрепарата
в дозе 0,75 мл/гол.
Экономическая эффективность использования пробиотика. Результаты экономической эффективности использования пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах кросса Кобб 500 представлены в таблице 99.
В результате проведенной предварительной экономической эффективности применения в рационе цыплят-бройлеров кросса Кобб 500 микробной добавки Галлобакт-Ф-Плюс выявлено, что прибыль в 1–4-й опытных группах составила 3 481,94; 3 652,95; 3 870,26 и 3 764,24 руб., против 3 261,23 руб. в контрольной группе, что выше на 220,71; 391,72; 604,03 и 503,01 руб., соответственно. В целом, наилучшие результаты были получены в 3-й опытной группе, в которой экономический эффект превышал контрольную группу на 18,67 %.
Таким образом, экономически целесообразно проводить производственную апробацию пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах кросса Кобб 500 в дозе, составляющей 0,75 мл/гол.
Таблица 99 – Экономическая эффективность использования пробиотика Галлобакт-Ф-Плюс
| Показатель | Группа | |||||
| контрольная | 1-я опытная | 2-я опытная | 3-я опытная | 4-я опытная | ||
| Поголовье в начале опыта, гол. | 60 | 60 | 60 | 60 | 60 | |
| Сохранность, % | 95,0 | 98,3 | 100 | 100 | 100 | |
| Поголовье в конце опыта, гол. | 57 | 59 | 60 | 60 | 60 | |
| Масса тушки на реализацию | ||||||
| Масса потрошенной тушки, г | 1810,927,54 | 1835,837,31 | 1860,537,45* | 1898,797,42* | 1882,387,37* | |
| Масса потрошенной тушки всего, кг | 103,22 | 108,31 | 111,63 | 113,93 | 112,94 | |
| Затраты комбикорма за период выращивания | ||||||
| На голову, г | 4587,81 | 4608,48 | 4625,78 | 4628,59 | 4627,93 | |
| На все поголовье, кг | 261,51 | 271,90 | 277,55 | 277,72 | 277,68 | |
| Затраты пробиотика | ||||||
| Всего, мл | – | 132,75 | 270,0 | 405,0 | 540,0 | |
| Экономическая эффективность | ||||||
| Цена 1 кг комбикорма, руб. | 27 | |||||
| Стоимость израсходованного корма всего, руб. |
7 060,77 | 7 341,30 | 7 493,85 | 7 498,44 | 7 497,36 | |
| Цена 1 л пробиотика, руб. | – | 60 | ||||
| Стоимость пробиотика всего, руб. | – | 7,76 | 16,20 | 24,30 | 32,40 | |
| Стоимость израсходованного корма и пробиотика, руб. | 7 060,77 | 7 349,06 | 7 510,05 | 7 522,74 | 7 529,76 | |
| Цена реализации 1 кг мяса птицы, руб. | 100,0 | |||||
| Выручка от реализации мяса птицы, руб. | 10 322,00 | 10 831,00 | 11 163,00 | 11 393,00 | 11 294,00 | |
| Прибыль от реализации мяса птицы, руб. | 3 261,23 | 3 481,94 | 3 652,95 | 3 870,26 | 3 764,24 | |
| Экономический эффект от использования пробиотика к контролю,руб.% |
–
– |
220,71
6,77 |
391,72
12,01 |
604,03
18,67 |
503,01
15,42 |
|
3 ПРОИЗВОДСТВЕННЫЕ ИСПЫТАНИЯ
Производственную апробацию эффективности применения разработанного пробиотика осуществляли в крестьянско-фермерских хозяйствах Краснодарского края и Республики Адыгея.
Эффективность использования пробиотика на перепелах. Схема производственных испытаний пробиотика Галлобакт-Ф-Плюс на перепелах представлена в таблице 100.
Таблица 100 – Схема производственных испытаний на перепелах при
использовании пробиотика Галлобакт-Ф-Плюс
| Группа | Условия кормления и выпаивания | Схема выпаивания, дни |
| Контрольная | Стандартный рацион (СР) и питьевая вода (ПВ) | |
| Опытная | СР, ПВ + 0,5 мл/гол. Галлобакт-Ф-Плюс | 1; 3; 5; 7; 14; 21; 28; 35; 42; 49 |
Согласно производственной схеме было сформировано две группы перепелов: контрольная группа – перепела получали стандартный рацион и имели свободный доступ к воде; опытная группа – перепела получали стандартный рацион, а также дополнительно в воду вводился пробиотик Галлобакт-Ф-Плюс в дозе 0,5 мл/гол. Продолжительность производственной апробации ‒ 56 дней. Результаты производственной апробации на перепелах Техасской породы продемонстрированы в таблице 101.
Таблица 101 – Производственная эффективность использования пробиотика Галлобакт-Ф-Плюс на перепелах Техасской породы
| Показатель | Группа | |
| контрольная | опытная | |
| 1 | 2 | 3 |
| Поголовье в начале испытаний, гол. | 2 000 | 2 000 |
| Сохранность, % | 95,6 | 99,0 |
| Поголовье в конце испытаний, гол. | 1912 | 1980 |
| Динамика живой массы, г. | ||
| Суточные | 10,74±0,34 | 10,69±0,28 |
| 56-е сутки | 321,23±2,01 | 346,29±2,05* |
| Масса тушки на реализацию | ||
| Потрошенная тушка, г | 236,67±1,65 | 262,59±1,69* |
| Всего, кг | 452,50 | 519,92 |
| Продолжение таблицы 101 | ||
| 1 | 2 | 3 |
| Затраты комбикорма за период выращивания | ||
| На голову, г | 1059,23 | 1098,47 |
| На все поголовье, кг | 2 025,24 | 2 174,96 |
| Затраты пробиотика | ||
| Пробиотика всего, мл | – | 9 900,0 |
| Экономическая эффективность | ||
| Цена 1 кг комбикорма, руб. | 30 | |
| Стоимость израсходованного корма всего, руб. | 60 757,20 | 65 248,80 |
| Цена 1 л пробиотика, руб. | – | 60 |
| Стоимость пробиотика, руб. | – | 594,00 |
| Стоимость израсходованного корма и добавки, руб. |
60 757,20 | 65 842,80 |
| Цена реализации 1 кг мяса птицы, руб. | 400,0 | |
| Выручка от реализации мяса птицы, руб. | 181 000,00 | 207 968,00 |
| Прибыль от реализации мяса птицы, руб. | 120 242,80 | 142 125,20 |
| Экономический эффект от использования добавки к контролю,руб.% |
–
– |
21 884,40
18,19 |
| Рентабельность затрат, % | 197,91 | 215,86 |
| к контролю, % | – | 17,95 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||
В результате проведенных производственных испытаний на перепелах Техасской породы установлено, что применение разработанного пробиотика
в опытной группе способствовало повышению жизнеспособности птицы, что отразилось на показателе сохранности, который был выше, чем в группе контроля на 2,4 %. Выявлена статистически достоверная разница в живой массе опытной группы по сравнению с контрольной, которыя была выше на 7,8 % (P < 0,05).
При расчете экономических показателей установлено, что прибыль
в опытной группе составила 142 125,20 руб., что на 14,35 % оказалось выше, чем в контрольной группе. Рентабельность затрат в опытной группе была выше, чем в контрольной группе на 17,95 %.
Результаты производственной апробации на перепелах породы Фараон представлены в таблице 102.
В результате проведенных производственных испытаний на перепелах породы Фараон установлено, что применение разработанного биопрепарата
в опытной группе способствовало повышению жизнеспособности птицы, что отразилось на показателе сохранности, который был выше, чем в группе контроля на 3,6 %. Выявлена статистически достоверная разница в живой массе опытной группы по сравнению с контрольной, которыя была выше на 7,2 % (P < 0,05). Также выявлена статистически достоверная разница по массе потрошенной тушки в опытной группе по сравнению с контрольной, которая составила в опытной группе 234,61 г, что на 7,4 % выше, чем в группе контроля (P < 0,05). В целом, экономический эффект от применения пробиотика Галлобакт-Ф-Плюс в опытной группе был выше, чем в контрольной, на 18,1 %. Рентабельность затрат в опытной группе была выше, чем в контрольной группе
на 18,43 %.
Таблица 102 – Производственная эффективность использования пробиотика Галлобакт-Ф-Плюс на перепелах породы Фараон
| Показатель | Группа | |
| контрольная | опытная | |
| Поголовье в начале испытаний, гол. | 1 000 | 1 000 |
| Сохранность, % | 95,1 | 98,7 |
| Поголовье в конце испытаний, гол. | 951 | 987 |
| Динамика живой массы, г. | ||
| Суточные | 8,83±0,15 | 8,85±0,19 |
| 56-е сутки | 293,57±1,87 | 314,72±1,81* |
| Масса тушки на реализацию | ||
| Потрошенная тушка, г | 212,38±1,77 | 234,61±1,70* |
| Всего, кг | 201,97 | 231,56 |
| Затраты комбикорма за период выращивания | ||
| На голову, г | 959,14 | 987,29 |
| На все поголовье, кг | 912,14 | 974,46 |
| Затраты пробиотика | ||
| Пробиотика всего, мл | – | 4 935,0 |
| Экономическая эффективность | ||
| Цена 1 кг комбикорма, руб. | 30 | |
| Стоимость израсходованного корма всего, руб. | 27 364,20 | 29 233,80 |
| Цена 1 л пробиотика, руб. | – | 60 |
| Стоимость пробиотика, руб. | – | 296,10 |
| Стоимость израсходованного корма и добавки, руб. | 27 364,20 | 29 529,90 |
| Цена реализации 1 кг мяса птицы, руб. | 400,0 | |
| Выручка от реализации мяса птицы, руб. | 80 788,00 | 92 624,00 |
| Прибыль от реализации мяса птицы, руб. | 53 423,80 | 63 094,10 |
| Экономический эффект от использования добавки к контролю,руб.% |
–
– |
9 670,30
18,1 |
| Продолжение таблицы 102 | ||
| 1 | 2 | 3 |
| Рентабельность затрат, % | 195,23 | 213,66 |
| к контролю, % | – | 18,43 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||
Таким образом, применение пробиотика Галлобакт-Ф-Плюс в дозе 0,5 мл/гол. способствуют повышению экономической эффективности и рентабельности выращивания перепелов мясной продуктивности.
Эффективность использования пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах быстрорастущих кроссов. Схема производственных испытаний представлена в таблице 103.
Таблица 103 – Схема производственных испытаний на цыплятах-бройлерах быстрорастущих кроссов при использовании пробиотика
Галлобакт-Ф-Плюс
| Группа | Условия кормления и выпаивания | Схема выпаивания, дни |
| Контрольная | Стандартный рацион (СР) и питьевая вода (ПВ) | |
| Опытная | СР, ПВ + 0,75 мл/гол Галлобакт-Ф-Плюс | 1; 3; 5; 7; 14; 21; 28; 35 |
Согласно производственной схеме было сформировано две группы птиц: контрольная группа – цыплята-бройлеры получали стандартный рацион и имела свободный доступ к воде; опытная группа – птица получала стандартный рацион, а также дополнительно в воду вводился пробиотик Галлобакт-Ф-Плюс
в дозе 0,75 мл/гол. Продолжительность производственной апробации ‒ 42 дня. Результаты производственной апробации на цыплят-бройлеров кросса Росс 308 представлены в таблице 104.
Таблица 104 – Производственная эффективность использования пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах кросса Росс 308
| Показатель | Группа | |
| контрольная | опытная | |
| Поголовье в начале испытаний, гол. | 500 | 500 |
| Сохранность, % | 96,2 | 98,8 |
| Поголовье в конце испытаний, гол. | 481 | 494 |
| Динамика живой массы, г. | ||
| Суточные | 40,87±0,39 | 41,05±0,54 |
| 42-е сутки | 2672,36±7,02 | 2755,76±6,87* |
| Продолжение таблицы 104 | ||
| 1 | 2 | 3 |
| Масса тушки на реализацию | ||
| Потрошенная тушка, г | 1806,74±8,94 | 1914,38±8,78* |
| Всего, кг | 869,94 | 945,71 |
| Затраты комбикорма за период выращивания | ||
| На голову, г | 4787,52 | 4841,87 |
| На все поголовье, кг | 2 305,19 | 2 391,88 |
| Затраты пробиотика | ||
| Пробиотика всего, мл | – | 3 334,5 |
| Экономическая эффективность | ||
| Цена 1 кг комбикорма, руб. | 27 | |
| Стоимость израсходованного корма всего, руб. |
62 240,13 | 64 580,90 |
| Цена 1 л пробиотика, руб. | – | 60 |
| Стоимость пробиотика, руб. | – | 200,9 |
| Стоимость израсходованного корма и пробиотика, руб. |
62 240,13 | 64 780,99 |
| Цена реализации 1 кг мяса птицы, руб. | 100 | |
| Выручка от реализации мяса птицы, руб. | 86 994,50 | 94 570,50 |
| Прибыль от реализации мяса птицы, руб. | 24 754,37 | 29 789,51 |
| Экономический эффект от использования пробиотика к контролю,
руб. % |
–
– |
5 035,14
20,34 |
| Рентабельность затрат, % | 39,77 | 45,99 |
| к контролю, % | – | 6,22 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||
В результате проведенных производственных испытаний на цыплятах-бройлерах кросса Росс 308 установлено, что применение пробиотика Галлобакт-Ф-Плюс в опытной группе способствовало повышению жизнеспособности птицы, что отразилось на показателе сохранности, который был выше, чем
в группе контроля, на 2,5 %. Выявлена статистически достоверная разница
в живой массе опытной группы по сравнению с контрольной, которыя была выше на 3,1 % (P < 0,05). При расчете экономических показателей установлено, что прибыль в опытной группе составила 29 789,51 руб., что на 20,34 % оказалось выше, чем в контрольной группе. Рентабельность затрат в опытной группе была выше на 6,22 %.
Результаты производственной апробации на цыплят-бройлеров кросса Кобб 500 представлены в таблице 105.
Таблица 105 – Производственная эффективность использования пробиотика Галлобакт-Ф-Плюс на цыплятах-бройлерах кросса Кобб 500
| Показатель | Группа | |
| контрольная | опытная | |
| Поголовье в начале испытаний, гол. | 750 | 750 |
| Сохранность, % | 97,1 | 98,9 |
| Поголовье в конце испытаний, гол. | 728 | 742 |
| Динамика живой массы, г. | ||
| Суточные | 42,54±0,33 | 42,21±0,40 |
| 42-е сутки | 2610,52±5,77 | 2693,73±6,02* |
| Масса тушки на реализацию | ||
| Потрошенная тушка, г | 1808,667,22 | 1899,197,30* |
| Всего, кг | 1 317,16 | 1 408,73 |
| Затраты комбикорма за период выращивания | ||
| На голову, г | 4581,49 | 4625,87 |
| На все поголовье, кг | 3 336,47 | 3 431,22 |
| Затраты пробиотика | ||
| Пробиотика всего, мл | – | 5 006,7 |
| Экономическая эффективность | ||
| Цена 1 кг комбикорма, руб. | 27 | |
| Стоимость израсходованного корма всего, руб. |
90 084,76 | 92 642,94 |
| Цена 1 л пробиотика, руб. | – | 60 |
| Стоимость пробиотика, руб. | – | 300,41 |
| Стоимость израсходованного корма и пробиотика, руб. |
90 084,76 | 92 943,35 |
| Цена реализации 1 кг мяса птицы, руб. | 100 | |
| Выручка от реализации мяса птицы, руб. | 131 716,00 | 140 873,00 |
| Прибыль от реализации мяса птицы, руб. | 41 631,24 | 47 929,65 |
| Экономический эффект от использования пробиотика к контролю,
руб. % |
–
– |
6 298,41
15,13 |
| Рентабельность затрат, % | 46,21 | 51,57 |
| к контролю, % | – | 5,36 |
| Примечание: * – разница с контролем достоверна (P < 0,05). | ||
В результате проведенных производственных испытаний на цыплятах-бройлерах кросса Кобб 500 установлено, что применение пробиотика Галлобакт-Ф-Плюс в опытной группе способствовало повышению прибыли в опытной группе, которая составила 47 929,65 руб., что на 6 298,41 руб. или 15,13 % выше, чем в контрольной группе. Рентабельность затрат в опытной группе была выше на 5,36 %.
Таким образом, применение пробиотика Галлобакт-Ф-Плюс в дозе 0,75 мл/гол. на цыплятах-бройлерах быстрорастущих кроссов, в частности Росс 308 и Кобб 500, способствует повышению экономических показателей при выращивании данного вида птицы.
ЗАКЛЮЧЕНИЕ
На основании проведенных исследований можно сделать следующие выводы:
- Получены чистые культуры нескольких кисломолочных штаммов бактерий, выделенных из автохтонных эволюционно закрепленных микробных сообществ ЖКТ диких куриных Краснодарского края. В частности культуры штаммов Lactobacillus salivarius 16РС20, Lactobacillus curvatus 13PC723
и Lactobacillus sp. 100PC13. - Созданы геномные библиотеки для высокоэффективного секвенирования с использованием набора Nextera DNA Flex Library Prep компании Illumina для осуществления высокоэффективного полногеномного секвенирования на секвенаторе нового поколения Illumina MiSeq.
- Получены полногеномные сиквенсы пробиотических штаммов бактерий, принадлежащих к видам Lactobacillus salivarius, Lactobacillus curvatus, Lactobacillus parabuchneri и Lactobacillus heilongjiangensis. При этом секвенирование некоторых штаммов было выполнено несколько раз.
- Полногеномные последовательности подготовлены к депонированию
и размещены в международной базе NCBI (англ. National Center for Biotechnological Information, Национальный центр биотехнологической информации США) с присвоением соответствующих индивидуальных номеров: Lactobacillus salivarius PC20b (VRA_1sq_f) (Assembly: GCA_009683065.1), Lactobacillus salivarius PC20b (VRA_01-1sq_f) (Assembly: GCA_009683115.1), Lactobacillus curvatus PC723_5 (VRA_2sq_f) (Assembly: GCA_009683015.1), Lactobacillus curvatus PC723_5 (VRA_2sq_n) (Assembly: GCA_009683145.1). - Идентифицированы последовательности девяти бактериоцинов и осуществлен их биоинформационный анализ.
- Использование в составе рецептуры пробиотика Галлобакт-Ф-Плюс полисахаридной смеси, в частности альгината натрия в дозе 0,3 %, способствует высокой резистентности выделенных лактобацил к фенолу, желчи, хлориду натрия и желудочному соку, что способствует максимальной приживаемости культур в желудочно-кишечном тракте птиц.
- Выделенные лактобациллы обладают свойствами, характерными для молочнокислых бактерий. Они проявляют высокую кислотообразующую способность, способны утилизировать широкий спектр сахаров, что дает им преимущества в конкуренции за субстрат.
- Многократный пассаж отобранных культур с терапевтическими дозами антибиотиков позволило повысить устойчивость лактобактерий к ним; по антагонистическим свойствам штаммы обладаот угнетающим действием на полевую и эталонную патогенную микрофлору без ингибирования роста представителей нормофлоры кишечника; по антиадгезивным свойствам исследуемые культуры обладают положительным действием на адгезию E. coli и S. aureus
к эритроцитам крови барана; по адгезивным характеристикам лактокультуры обладают высокоадгезивной активностью по отношению к энтероцитам кишечника куриного эмбриона (коэффициент прикрепительной способности 12,417,5 ед.). - При изучении безопасности лактобактерий установлено, что исследуемые микроорганизмы не продуцируют гемолизин (результат гемолиза – отрицательный), а при даче лактобактерий мышам, не было зафиксировано гибели и заболевания животных. Изучаемые штаммы микроорганизмов безопасны и могут быть использованы в качестве компонентов для пробиотических добавок.
- Пробиотик Галлобакт-Ф-Плюс малотоксичен для животных, не вызывает их гибели, как в остром, подостром, так и в хроническом экспериментах, что по ГОСТ 12.1.007-76 «Вредные вещества» позволяет отнести его к 4-му классу опасности. Длительное применение пробиотика не оказывает отрицательного воздействия на общее состояние лабораторных животных, на процессы пищеварения, морфологические и биохимические показатели крови. Галлобакт-Ф-Плюс не проявляет раздражающего действия на организм подопытных животных.
- Изучение лечебно-профилактического действия пробиотика Галлобакт-Ф-Плюс на модели экспериментального дисбактериоза в кишечнике крыс показало, что биопрепарат способствует повышению концентрации представителей нормофлоры (лакто- и бифидобактерий) и снижает численность представителей патогенной и условно-патогенной микробиоты, с последующим достижением показателей характерных для здоровых животных.
- Изучение фармакологических свойств пробиотической добавки Галлобакт-Ф-Плюс на организм перепелов (в дозировке 0,5 мл/гол.) и цыплят-бройлеров (в дозировке 0,75 мл/гол.) мясной продуктивности способствовало следующим изменениям:
- наблюдалось повышение сохранности: перепелов – на 3,5 % (Техасская порода) и 5,0 % (порода Фараон), цыплят-бройлеров – на 6,7 % (кросс Росс 308) и 5,0 % (кросс Кобб 500), живой массы: перепелов – на 6,3 % (Техасская порода) и 7,2 % (порода Фараон), цыплят-бройлеров – на 2,8 % (кросс Росс 308)
и 3,1 % (кросс Кобб 500), прироста: перепелов – на 6,6 % (Техасская порода)
и 7,4 % (порода Фараон), цыплят-бройлеров – на 2,9 % (кросс Росс 308) и 3,1 % (кросс Кобб 500), снижение конверсии комбикорма: у перепелов – на 1,5 % (Техасская порода) и 1,8 % (порода Фараон), цыплят-бройлеров – на 2,2 % (кросс Росс 308) и 1,4 % (кросс Кобб 500); - применение пробиотика способствовало интенсификации обменных процессор организма птицы за счет усиления гемо- и эритропоэза, повышается в крови перепелов количество эритроцитов и гемоглобина на 3,7 и 3,8 % (Техасская порода), 4,9 и 3,4 % (порода Фараон), у цыплят-бройлеров – на 8,1
и 4,7 % (кросс Росс 308), 8,2 и 4,6 % (кросс Кобб 500); стимулирует обмен белков, который характеризуется повышением содержания в сыворотки крови перепелов общего белка и белкового коэффициента (А/Г) на 10,6 и 9,1 % (Техасская порода), 9,3 и 7,9 % (порода Фараон), у цыплят-бройлеров – на 4,6 и 8,2 % (кросс Росс 308), 6,3 и 12,3 % (кросс Кобб 500); обмен жиров – за счет снижением количества холестерина в сыворотки крови перепелов на 5,7 % (Техасская порода) и 7,8 % (порода Фараон), у цыплят-бройлеров – на 9,3 % (кросс Росс 308) и 4,9 % (кросс Кобб 500); обмен минеральных веществ – за счет повышения уровня фосфора и кальция в сыворотки крови перепелов на 4,1 и 7,8 % (Техасская порода), 7,1 и 16,5 % (порода Фараон), у цыплят-бройлеров – на 2,8
и 4,8 % (кросс Росс 308), 3,5 и 2,8 % (кросс Кобб 500); повышается уровень неспецифической резистентности организма птиц за счет достоверного увеличения бактерицидной и лизоцимной активностей сыворотки крови у перепелов на 9,2 и 6,0 % (Техасская порода), 10,4 и 4,0 % (порода Фараон), у цыплят-бройлеров – на 4,5 и 3,6 % (кросс Росс 308), 2,8 и 2,6 % (кросс Кобб 500); - наблюдается положительное влияние на процессы пищеварения, о чем свидетельствует повышение переваримости органического вещества, сырого протеина, сырого жира и клетчатки у перепелов на 6,7; 5,9; 4,9 и 4,8 % (Техасская порода), 6,4; 4,1; 5,2 и 1,8 % (порода Фараон), у цыплят-бройлеров – на 2,4; 5,0; 1,4 и 0,9 % (кросс Росс 308), 3,3; 5,8; 2,3 и 2,3 % (кросс Кобб 500), соответственно; пробиотик стимулирует жизнедеятельность нормофлоры в кишечнике птиц, о чем свидетельствует увеличение количества в их химусе молочнокислой микрофлоры и бифидобактерий у перепелов в 3,5 и 3,0 раза (Техасская порода), 2,2 и 1,5 раза (порода Фараон), у цыплят-бройлеров – на 1,9 и 1,3 раза (кросс Росс 308), 1,5 и 1,3 раза (кросс Кобб 500) при одновременном снижении уровня условно-патогенной микробиоты;
- повышается мясная продуктивность и качество продукции птицеводства, а именно увеличивается масса потрошенной тушки у перепелов на 6,3 % (Техасская порода) и 7,1 % (порода Фараон), у цыплят-бройлеров – на 5,5 % (кросс Росс 308) и 4,8 % (кросс Кобб 500); повышается содержание в мышечной ткани белка и снижается уровень жира у перепелов на 0,92 и 0,52 % (Техасская порода) и 0,77 и 0,49 % (порода Фараон), у цыплят-бройлеров – на 0,3 и 0,2 % (кросс Росс 308) и 0,63 и 0,31 % (кросс Кобб 500), соответственно, при одновременном улучшении аминокислотного скора; мясо, полученное от опытных групп птиц, может быть использовано в пищу независимо от дозы применения пробиотика; выявлено положительное влияние пробиотика на массу иммунокомпетентных органов цыплят-бройлеров быстрорастущего кросса.
- Результаты производственных испытаний показали, что применение
в рационе перепелов и цыплят-бройлеров пробиотика Трилактокор по отношению к контролю экономически эффективно:
- использование пробиотика Трилактокор в дозе 0,5 мл/гол. в системе поения перепелов способствовало повышению экономической эффективности на 18,2 %, а рентабельности затрат – на 18,0–18,4 %;
- применение пробиотика Трилактокор в дозе 0,75 мл/гол. в рационе цыплят-бройлеров быстрорастущих кроссов способствовало повышению экономической эффективности на 13,2‒20,3 %, а рентабельности затрат – на 4,6‒6,2 %.
Исходя из полученных в ходе научно-хозяйственных исследований результатов, были сделаны следующие предложения производству:
- Для повышения сохранности, продуктивности перепелов мясных пород, снижения расхода корма и повышения рентабельности, рекомендуется использовать путем выпаивания птице жидкую пробиотическую добавку Галлобакт-Ф-Плюс в дозе 0,5 мл/гол. согласно схеме (1-е, 3-е, 5-е, 7-е сутки и далее один раз в неделю до реализации).
- Для повышения сохранности, продуктивности цыплят-бройлеров, снижения расхода кормов и повышения рентабельности, рекомендуется выпаивать птице жидкую пробиотическую добавку Галлобакт-Ф-Плюс в дозе 0,75 мл/гол. согласно схеме ‒ 1-е, 3-е, 5-е, 7-е сутки и далее один раз в неделю до реализации.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
- Андрейчин, М. А. Антимикробные свойства желчи и желчных кислот / М. А. Андрейчин // Антибиотики. – 1980. – № 25(12). – С. 936–939.
- Балтрашевич, А. К. Способ определения чувствительности, анаэробных микроорганизмов к антибиотикам с использованием стандартных дисков / А. К. Балтрашевич, Т. П. Комаровская // Антибиотики. – 1982. – № 8. – С. 32–36.
- Водоросли – источник биополимеров, биологически активных веществ и субстрат в биотехнологии. Ч. 1. Биополимеры клеток тканей водорослей / И. А. Наумов [и др.] // Вестник Казанского технологического университета. ‒ 2014. ‒ Т. 17, № 1. ‒ С. 188–193.
- Воробьёва, Л. И. Промышленная микробиология / Л. И. Воробьёва. – М. : Изд-во МГУ, 1989. – 246 с.
- Выделение и отбор бактерий рода Lactobacillus – основы пробиотических препаратов / И. А. Буряко [и др.] // Пробиотики, пребиотики и синбиотики и функциональные продукты питания : материалы Междунар. конф. – М., 2004. – С. 19.
- Гвоздяк, Р. И. Микробный полисахарид ксантан / Р. И. Гвоздяк, М. С. Матышевская. Киев : Наукова думка, 1989. – 212 с.
- Глушанова, Н. А. Биологические свойства лактобацилл / Н. А. Глушанова // Бюл. сиб. медицины. – 2003. – № 4. – С. 50–58.
- Горощенко, Г. Российский рынок йогурта / Г. Горощенко // Молочная промышленность. 2003. – № 10. – С. 57.
- ГОСТ 10444.11-89. Продукты пищевые. Методы определения молочнокислых микроорганизмов. – Введ. 01.01.1994. – М. : Госстандарт России : Изд-во стандартов, 2010. – 15 с.
- ГОСТ 12.1.007-76. Система стандартов безопасности труда (ССБТ). Вредные вещества. Классификация и общие требования безопасности. – М. : Стандартинформ, 2007. – 7 с.
- ГОСТ 13496.15-97. Корма, комбикорма, комбикормовое сырье. Метод определения содержания сырого жира. – Введ. 1999-01-01. – М. : Изд-во стандартов, 2005. – 13 с.
- ГОСТ 13496.2-91. Корма, комбикорма, комбикормовое сырье. Метод определения сырой клетчатки. – Введ. 1992-07-01. – М. : Изд-во стандартов, 2002. – 6 с.
- ГОСТ 13496.3-92. Корма, комбикорма, комбикормовое сырье. Метод определения содержания влаги. – Введ. 1993-01-01. – М. : Изд-во стандартов, 2002. – 4 с.
- ГОСТ 13496.4-93. Корма, комбикорма, комбикормовое сырье. Метод определения содержания азота и сырого протеина. – Введ. 1995-01-01. – М. : Изд-во стандартов, 2002. – 17 с.
- ГОСТ 23042-78. Мясо и мясные продукты. Метод определения жира. – Введ. 1988-01-01. – М. : Изд-во стандартов, 2010. – 5 с.
- ГОСТ 25011-81. Мясо и мясные продукты. Метод определения белка. – Введ. 1983-01-01. – М. : Изд-во стандартов, 2010. – 7 с.
- ГОСТ 26226-95. Корма, комбикорма, комбикормовое сырье. Метод определения сырой золы. – Введ. 2003-07-01. – М. : Изд-во стандартов, 2010. – 8 с.
- ГОСТ 26712-94. Удобрения органические. Общие требования к методам анализа. – Введ. 01.01.1996. – М. : Госстандарт России : Изд-во стандартов, 1996. – 10 с.
- ГОСТ 26713-85. Удобрения органические. Метод определения влаги
и сухого остатка. – Введ. 19.12.1985. – М. : Госстандарт Союза ССР : Изд-во стандартов, 1985. – 7 с. - ГОСТ 31962-2013. Мясо кур (тушки кур, цыплят, цыплят-бройлеров
и их части). Технические условия. – Введ. 2014-07-01. – М. : Изд-во стандартов, 2016. – 12 с. - ГОСТ 9225-84. Молоко и молочные продукты. Методы микробиологического анализа [текст]. – Введ. 1986–01–01. – М. : Стандартинформ, 2009. – 15 с.
- ГОСТ 9793-74. Мясные продукты. Метод определения содержания влаги. – Введ. 1975-01-01. – М. : Изд-во стандартов, 2010. – 4 с.
- ГОСТ Р 50396.1-92. Мясо птицы, субпродукты и полуфабрикаты птичьи. Метод определения количества мезофильных аэробных и факультативно-анаэробных микроорганизмов. – Введ. 1994-01-01. – М. : Госстандарт России, 1993. – 5 с.
- ГОСТ Р 51331-99. Продукты молочные. Йогурты. Общие технические условия. – Введ.19.10.1999. – М. : Стандартинформ, 2008. – 22 с.
- ГОСТ Р 51478-99. Мясо и мясные продукты. Контрольный метод определения концентрации водородных ионов (pH). – Введ. 2001-01-01. – М. : Изд-во стандартов, 2010. – 5 с.
- ГОСТ Р 51944-2002. Мясо птицы. Методы определения органолептических показателей, температуры и массы. – Введ. 2003-07-01. – М. : Изд-во стандартов, 2010. – 8 с.
- ГОСТ Р 54673-2011. Мясо перепелов (тушки). Технические условия. – Введ. 2013-01-01. – М. : Изд-во стандартов, 2013. – 12 с.
- ГОСТ Р ИСО 10993.10-99. Изделия медицинские. Оценка биологического действия медицинских изделий. Часть 10. Исследование раздражающего и сенсибилизирующего действия. – Введ. 1999-12-29. – М. : Госстандарт России, 2000. – 38 с.
- ГОСТ Р ИСО 10993-11-2009. Изделия медицинские. Оценка биологического действия медицинских изделий. Часть 11. Исследования общетоксического действия. – Введ. 2009-10-20. – М. : Стандартинформ, 2010. – 27 с.
- Губанова, Т. С. Йогурт: потребители и предпочтения / Т. С. Губанова // Молочная промышленность. 2001. № 10. – С. 89.
- Ермоленко, Е. И. Бактериоцины энтерококков: проблемы и перспективы использования / Е. И. Ермоленко // Вестник Санкт-Петербургского университета. Серия 11. – 2009. – Вып. 3.
- Забодалова, Л. А. Биотехнология комбинированных молочных продуктов с использованием компонентов сои : автореф. дисс. … д-ра тех. наук : 05.18.04 / Забодалова Людмила Александровна. Кемерово, 2000. – 36 с.
- Зельцер, И. З. О прогнозировании дисбиотического эффекта при токсико-гигиенической оценке антибиотиков / И. З. Зельцер // Антибиотики и химиотерапия. – 1989. – № 6. – С. 440–442.
- Исследование ферментативных свойств кормовых добавок / А. Н. Швыдков, А. Е. Мартышенко, Н. Н. Ланцева, В. П. Чебаков, Л. А. Кобцева // Успехи современного естествознания. – 2014. – № 11-2. – С. 49–53.
- Кощаев, А. Г. Биотехнология производства и применение функциональных кормовых добавок для птицы : дисс. … д-ра биол. наук : 16.00.04 / Кощаев Андрей Георгиевич. – 2008. – 425 с.
- Кудрявцев, А. А. Клиническая гематология животных / А. А. Кудрявцев, Л. А. Кудрявцева. – М. : Колос, 1973. – 376 с.
- Лабинская, А. С. Микробиология с техникой микробиологических исследований / А. С. Лабинская. – М. : Медицина, 1978. – 394 с.
- Липатов, Н. Н. Молочная промышленность в XXI веке / Н. Н. Липатов // Пищевая и перерабатывающая промышленность. 1987. № 10. – С. 4.
- Мартынов, А. В. Потенциал молочной продукции пониженной жирности / А. В. Мартынов // Молочная промышленность. 1999. № 8. – С. 1112.
- Методика проведения научных и производственных исследований по кормлению сельскохозяйственной птицы : рекомендации / Ш. А. Имангулов, И. А. Егоров, Т. М. Околелова, А. Н. Тишенков. – Сергиев Посад : ВНИТИП, 2013. – 52 с.
- Митрохин, С. Д. Микробиологические и биохимические показатели изменения микробной экологии толстой кишки крыс под влиянием рифампицина / С. Д. Митрохин, Б. А. Шендеров // Антибиотики и химиотерапия. ‒ 1989. ‒ Т. 34, № 6. ‒ С. 448‒452.
- МУ 2.3.2.2789-10. Методические указания по санитарно-эпидемиологической оценке безопасности и функционального потенциала пробиотических микроорганизмов, используемых для производства пищевых продуктов. − М. : Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2011. – 104 с.
- Назырова, Н. Р. Антибиотикорезистентность штаммов, входящих
в состав препаратов-пробиотиков / Н. Р. Назырова, Р. Х. Тимербаева, М. М. Туйгунов // Химия, химическая технология и биотехнология на рубеже тысячелетий : материалы IV Междунар. науч. конф. – Томск, 2006. – Т 2. – С. 436–438. - Назырова, Н. Р. Влияние экстрактов лекарственных растений на биологическую активность штамма Lactobacterium plantarum 8P-A3 : дисс. … канд. биол. наук : 03.00.07, 03.00.04 / Назырова Наиля Рамилевна. ‒ Уфа, 2007. ‒ 147 с.
- Нуртдинова, А. Н. Разработка и изучение биологических свойств комплексного препарата ‒ Бифидоспорина : дисс. … канд. мед. наук : 03.00.07 / Нуртдинова Айгуль Наилевна. ‒ Уфа, 2003. ‒ 110 с.
- Осипова, И. Г. Изучение эффективности пробиотиков на экспериментальных моделях / И. Г. Осипова, Е. А. Васильева // Совершенствование иммунобиологических средств профилактики, диагностики и лечения инфекционных болезней : материалы Всеросс. науч.-практ. конф. «Вакцинология 2006». ‒ М., 2006. ‒ С. 75‒76.
- Петров, Л. Н. Основы конструирования пробиотиков повышенной терапевтической активности / Л. Н. Петров // Пробиот. микроорганизмы – современное состояние вопроса и перспективы использования : тез. докл. Междунар. науч.-практ. конф. – М., 2002. – С. 12.
- Правила ветеринарного осмотра убойных животных и ветеринарно-санитарной экспертизы мяса и мясных продуктов: Ветеринарное законодательство. – Т. 4. – М. : Агропромиздат, 1988. – 38 с.
- Романчук, Л. А. Изучение влияния новых антимикробных препаратов, применяемых в комплексе с общей гнотобиологической изоляцией, на выжимаемость и микрофлору кишечника тотально облученных мышей / Л. А. Романчук, Д. А. Елизбаращвили, Г. А. Гинодман // ЖМЭИ. ‒ 1991. ‒ № 6. ‒ С. 4‒6.
- Смирнов, А. М. Научно-методологические аспекты исследования токсических свойств фармакологических лекарственных средств для животных / А. А. Смирнов, В. И. Дорожин. ‒ М. : Россельхозакадемия, 2008. – 120 с.
- Смирнова, Н. П. Определение бактерицидной активности сыворотки крови методом фотонефелометрии / Н. П. Смирнова, Т. А. Кузьмина // Журн. микробиологии. – 1966. – № 4. – С. 8–13.
- СП 2.2.1.3218-14. Санитарно-эпидемиологические правила. Санитарно-эпидемиологические требования к устройству, оборудованию и содержанию экспериментально-биологических клиник (вивариев). ‒ М., 2014. – 7 с.
- Способ определения активности лизоцима в слюне и сыворотке крови / В. И. Стогник [и др.] // Лаб. дело. – 1989. – № 8. – С. 54.
- Сультимова, Т. Д. Выделение активных бактериоцинобразующих лактококков и их практическое использование : автореф. дисс. … канд. биол. наук : 03.00.07 / Сультимова Татьяна Доржиевна. – М., 2006. – 30 с.
- Токсикологическая оценка новых препаратов для лечения и профилактики незаразных болезней животных : метод. указания / сост. А. И. Тишков, М. Н. Аргунов, Н. И. Ляшко. – Воронеж : ВНИИНБЖ, 1987. – 23 с.
- Фисинин, В. И. Использование нетрадиционных кормов в рационе птицы / В. И. Фисинин, И. А. Егоров, Т. Н. Ленкова // Птица и птицепродукты. – 2016. – № 4 – С. 1418.
- Хабриев, Р. У. Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ / Р. У. Хабриев. – М. : ОАО «Медицина», 2005. – 2-изд., перераб. и доп. – 832 с.
- Шацкая, Н. Г. Технологические аспекты производства молочных продуктов с низкими аллергическими свойствами и гипоаллергенных продуктов / Н. Г. Шацкая, П. Ф. Крашенинин, Г. Ю. Сажинов // Вопросы питания. 1994. № 1/2. – С. 49.
- Шендеров, Б. А. Медицинская микробная экология и функциональное питание. Т. I: Микрофлора человека и животных и ее функции / Б. А. Шендеров. – М. : ГРАНТЬ, 1998. 288 с.
- (Lactobacillus planetarium subsp. plantarum) Zhang-LL Bacteriocin and Nisin Composite Bacteriostatic Agent and its Use in Chilled Meat. (CN104531561 A) / Zhang, Hongxing, Liu, Hui, Bai, Yongqiang, Xu, Hongyuan, Duan, Huixia, Hao, Yanfang, Xie, Yuanhong, Jin, Junhua, Xiong, Lixia, Gao, Xiuzhi, 2015.
- ‘Normalization’ of germfree mice with anaerobically cultured caecal
flora of ‘normal’ mice / J. P. Koopman [et al.] // Lab Anim. – 1984. – Vol. 18. –
Р. 188–194. - 16s rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca / J. Gong
[et al.] // FEMS Microbiol Ecol. – 2007. – Vol. 59. – Р. 147–157. - A bacteriocin and food preservative, inhibits head and neck cancer tumorigenesis and prolongs survival / P. Kamarajan, T. Hayami, B. Matte, Y. Liu, T. Danciu, A. Ramamoorthy, F. Worden, S. Kapila, Y. Kapila, Z. P. Nisin // PLoS One. – 2015. – Vol. 10. – e0131008.
- A system for heterologous expression of bacteriocins in Lactobacillus sake / L. Axelsson, T. Katla, M. Bjørnslett, V. G. Eijsink, A. Holck // FEMS Microbiol. Lett. – 1998. – Vol. 168. – Р. 137–143.
- A tale of three next generation sequencing platforms: comparison of Ion Torrent, Pacific Biosciences and Illumina MiSeq sequencers / M. A. Quail [et al.] // BMC Genomics. – 2012. – Vol. 13. – Р. 341.
- Adeola, O. Opportunities and challenges in using exogenous enzymes to improve non-ruminant animal production / O. Adeola, A. J. Cowieson // J Animal Science. – 2011. – Vol. 89. – P. 3189–3218.
- Advances and needs for endotoxin-free production strains / S. Taguchi,
T. Ooi, K. Mizuno, H. Matsusaki // Appl Microbiol Biotechnol. – 2015. –
Vol. 99(22). – Р. 9349-9360. - Alloui, M. N. The usefulness of prebiotics and probiotics in modern poultry nutrition: a review / M. N. Alloui, W. Szczurek, S. Swiatkiewicz // Annals of Animal Science. – 2013. – Vol. 13. – P. 17–32.
- An antimicrobial peptide-3: effects on growth perfomance, nutrient retention, intestinal and faecal microflora and intestinal morphology of broilers /
S. C. Choi, S. L. Ingale, J. S. Kim, Y. K. Park, I. K. Kwon, B. J. Chae // British Poultry Science. – 2013. – Vol. 54. – P. 738–746. - Analysis, optimization and verification of Illumina-generated 16S rRNA gene amplicon surveys / M. C. Nelson, H. G. Morrison, J. Benjamino, S. L. Grim,
J. Graf // PLoS One. – 2014. – Vol. 9(4). – e94249. - Antibiotic alternatives: the substitution of antibiotics in animal husbanddry? / G. Cheng, H. Hao, S. Xie, X. Wang, M. Dai, L. Huang, Z. Yuan // Frontiers Microbiology. – 2014. – Vol. 5, аrticle 217.
- Antimicrobial activity of lactic acid bacteria against pathogenic and spoilage microorganism isolated from food and their control in wheat bread / D. Cizeikiene, G. Juodeikeine, A. Paskevicius, E. Bartkiene // Food Control. – 2013. –
Vol. 31. – Р. 539–545. - Apajalahti, J. Characteristics of the gastrointestinal microbial communities, with special reference to the chicken / J. Apajalahti, A. Kettunen, H. Graham // Worlds Poult Sci J. – 2004. – Vol. 60. – Р. 223–232.
- Bacteria within the gastrointestinal tract microbiota correlated with improved growth and feed conversion: Challenges presented for the identification of performance enhancing probiotic bacteria / D. Stanley, R. J. Hughes, M. S. Geier,
R. J. Moore // Front. Microbiol. – 2016. – Vol. 7. – Р. 187. - Bacteriocins from lactic acid bacteria: purification, properties and use as biopreservatives / J. L. Parada, C. R. Caron, A. B. P. Medeiros, C. R. Soccol // Braz. Arch. Biol. Technol. – 2007. – Р. 521–542.
- Bacteriocins of Lactobacillus gasseri K7 – Monitoring of gassericin K7 Aand B genes’ expression and isolation of an active component / A. Mavriˇc,
G. Tompa, A. Trmˇci’c, I. Rogelj, B. B. Matijaˇsi’c // Process Biochem. – 2014. – Vol. 49. – Р. 1251–1259. - Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria / R. Kumariya, A. K. Garsa,
Y. S. Rajput, S. K. Sood, N. Akhtar, S. Patel // Microb Pathog. – 2019. – Vol. 128. – Р. 171–177. - Bactofencin A, a new type of cationic bacteriocin with unusual immunity / E. F. O’Shea, P. M. O’Connor, O. O’Sullivan, P. D. Cotter, R. P. Ross, C. Hill //
mBio 4. – 2013. – e00498–e00513. - Beasley, S. Nisin-producing Lactococcus lactis strains isolated from
human milk / S. Beasley, P. Saris // Appl. Environ. Microbiol. – 2004. – Vol. 70. –
P. 5051–5053. - Bedford, M. R. Exogenous enzymes for pigs and poultry / M. R. Bedford, H. Schulze // Nutrition Research Reviews. – 1998. – Vol. 11. – P. 91–114.
- Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production /
N. Vieco-Saiz, Y. Belguesmia, R. Raspoet, E. Auclair, F. Gancel, I. Kempf,
D. Drider // Front Microbiol. – 2019. – Vol. 10. – Р. 57. - Bile acid metabolism by colonic bacteria in continuous culture: effect of starch and pH / S. U. Christl, H. P. Bartram, A. Paul, E. Kelber, W. Scheppach,
H. Kasper // Ann Nutrit Metabol. – 1997. – Vol. 41. – Р. 45–51. - Biodiversity of lactic acid bacteria / W. Liu, H. Pang, H. Zhang, Y. Cai // In: Lactic Acid Bacteria / Y. Zhang, Y. Cai (Eds.). – Springer Science +
Business Media, Dordrecht, The Netherlands, 2014. – Р. 103–203. – doi : https://doi.org/10.1007/978-94-017-8841-0_2. - Borda-Molina, D. Current perspective of the chicken gastrointestinal tract and its microbiome / D. Borda-Molina, J. Seifert, A. Camarinha-Silva // Comput Struct Biotechnol J. – 2018. – Vol. 16. – Р. 131–139.
- Borda-Molina, D. Current perspectives of the chicken gastrointestinal tract and its microbiome / D. Borda-Molina, J. Seifert, A. Camarinha-Silva // Comput. Struct. Biotechnol. J. – 2018. – Vol. 16. – Р. 131–139.
- Cellulose nanofiber (CNF)-sakacin-A active material: production, characterization and application in storage trials of smoked salmon / C. Mapelli, A. Musatti, A. Barbiroli, S. Saini, J. Bras, D. Cavicchioli, M. Rollini // J Sci Food Agric. – 2019, Aug 15. – Vol. 99(10). – Р. 4731–4738.
- Characteristics and genetic determinant of a hydrophobic peptide bacteriocin, carnobacteriocin A, produced by Carnobacterium piscicola LV17A / R. Worobo, T. Henkel, M. Sailer, K. Roy, J. Vederas, M. Stiles // Microbiology. – 1994. –
Vol. 140. – P. 517–526. - Characterization and heterologous expression of a class IIa bacteriocin, plantaricin 423 from Lactobacillus plantarum 423, in Saccharomyces cerevisiae /
C. A. Van Reenen, M. L. Chikindas, W. H. Van Zyl, L. M. Dicks // Int. J. Food Microbiol. – 2003. – Vol. 81. – Р. 29–40. - Characterization of leucocin A-UAL 187 and cloning of the bacteriocin gene from Leuconostoc gelidum / J. Hastings, M. Sailer, K. Johnson, K. Roy,
J. Vederas, M. Stiles // J. Bacteriol. – 1991. – Vol. 173. – P. 7491–7500. - Characterization of leucocin A-UAL 187 and cloning of the bacteriocin gene from Leuconostoc gelidum / J. W. Hastings, M. Sailer, K. Johnson, K. L. Roy, J. C. Vederas, M. E. Stiles // J. Bacteriol. – 1991. – Vol. 173. – Р. 7491–7500.
- Characterization of the mesB gene and expression of bacteriocins by Leuconostoc mesenteroides Y105 / Y.Hechard, J. M. Berjeaud, Y. Cenatiempo // Curr. Mi crobiol. – 1999. – Vol. 39. – P. 265–269.
- Characterization, production, and purification of leucocin H, a two-peptide bacteriocin from Leuconostoc MF215B / H. Blom, T. Katla, A. Holck, K. Sletten,
L. Axelsson, H. Holo // Curr. Microbiol. – 1999. – Vol. 39. – P. 43–48. - Characterizing and measuring bias in sequence data / M. G. Ross [et al.] // Gen Biol. – 2013. – Vol. 14. – R51.
- Chatterjee, M. Bacteriocin in harmony with ABC transporter exhibits antimicrobial activity / M. Chatterjee, A. Raichaudhuri // EC Microbiol. – 2017. –
Vol. 8. – Р. 3–10. - Chemical and genetic characterization of bacteriocins produced by Carnobacterium piscicola LV17B / L. Quadri, M. Sailers, K. L. Ron, J. C. Vederass,
M. Stiles // J. Biol. Chem. – 1994. – Vol. 269. – P. 12204–12211. - Chicken Gut Microbiota: Importance and Detection Technology // Front Vet Sci. – 2018. – Vol. 5. – Р. 254.
- Class IIa Bacteriocins: Current Knowledge and Perspectives / Y. Belguesmia, K. Naghmouchi, N.-E. Chihib, D. Drider. – 2011.
- Class III bacteriocin helveticin-M causes sublethal damage on target cells through impairment of cell wall and membrane / Z. Sun, X. Wang, X. Zhang, H. Wu, Y. Zou, P. Li, C. Sun, W. Xu, F. Liu, D. Wang // J. Ind. Microbiol. Biotechnol. – 2018. – Vol. 45.
- Classification of bacteriocins from grampositive bacteria / M. C. Rea,
R. P. Ross, P. D. Cotter, C. Hill // In: Prokaryotic Antimicrobial Peptides / D. Drider, S. Rebuffat (Eds.). – Springer, New York, NY, 2011. – Р. 29–53. - Cloning and expression of synthetic genes encoding the broad antimicrobial spectrum bacteriocins SRCAM 602, OR-7, E-760, and L-1077, by recombinant Pichia pastoris / S. Arbulu, J. J. Jiménez, L. Gútiez, L. M. Cintas, C. Herranz,
P. E. Hernández // BioMed. Res. Int. – 2015. – Vol. 1–11 (767183). - Cloning and heterologous production of Hiracin JM79, a Sec-Dependent bacteriocin produced by Enterococcus hirae DCH5, in lactic acid bacteria and Pichia pastoris / J. Sánchez, J. Borrero, B. Gómez-Sala, A. Basanta, C. Herranz,
L. M. Cintas, P. E. Hernández // Appl. Environ. Microbiol. – 2008. – Vol. 74. –
Р. 2471–2479. - Cloning strategies for heterologous expression of the bacteriocin enterocin A by Lactobacillus sakei Lb790, Lb. plantarum NC8 and Lb. casei CECT475 /
J. J. Jiménez, D. B. Diep, J. Borrero, L. Gútiez, S. Arbulu, I. F. Nes, C. Herranz,
L. M. Cintas, P. E. Hernández // Microb. Cell Factories. – 2015. – Vol. 14. – Р. 166. - Cloning, expression, and nucleotide sequence of genes involved in production of lactococcin DR, a bacteriocin from lactococcus lactis subsp. lactis /
A. Rince, S. Le Pogam, D. Thuault, C. M. Bourgeois, J. P. Le Pennec // Appl. Environ. Microbiol. – 1994. – Vol. 60. – P. 1652–1657. - Cloning, overexpression, purification of bacteriocin enterocin-B and structural analysis, interaction determination of enterocin-A, B against pathogenic bacteria and human cancer cells / D. Ankaiah, E. Palanichamy, C. B. Antonyraj,
R. Ayyanna, V. Perumal, S. B. Ahamed, V. Arul // Int J Biol Macromol. – 2018. – Vol. 116. – Р. 502–512. - Cloning, production, and functional expression of the bacteriocin enterocin A, produced by Enterococcus faecium T136, by the yeasts Pichia pastoris, Kluyveromyces lactis, Hansenula polymorpha, and Arxula adeninivorans / J. Borrero,
G. Kunze, J. J. Jiménez, E. Böer, L. Gútiez, C. Herranz, L. M. Cintas, P. E. Hernández // Appli. Environ. Microbiol. – 2012. – Vol. 78. – Р. 5956–5961. - Comparison of Next-Generation sequencing systems / L. Liu [et al.] //
J Biomed Biotechnol. – 2012. – 251364. - Comparison of three next-generation sequencing platforms for metagenomic sequencing and identification of pathogens in blood / K. G. Frey, J. E. Herrera-Galeano, C. L. Redden, T. V. Luu, S. L. Servetas, A. J. Mateczun,
V. P. Mokashi, K. A. Bishop-Lilly // BMC Genomics. – 2014. – Vol. 5. – Р. 96. - Cuozzo, S. Methods for the detection and concentration of bacteriocins produced by lactic acid bacteria / S. Cuozzo, F. Sesma // Food Microbiol. – 2001. – Vol. 14. – Р. 141–146.
- De Vuyst, L. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications / L. De Vuyst, F. Leroy // Mol. Microbiol. Biotechnol. – 2007. – Vol. 13. – P. 194–199.
- Deciphering chicken gut microbial dynamics based on high-throughput 16S rRNA metagenomics analyses / M. A. Shaufi, C. C. Sieo, C. W. Chong, H. M. Gan, Y. W. Ho // Gut Pathol. – 2015. – Vol. 7. – Р. 4.
- Development of the chick microbiome: how early exposure influences future microbial diversity / A. L. Ballou [et al.] // Front Veter Sci. – 2016. – Vol. 3. –
Р. 2. - Dibner, J. J. Antibiotic growth promoters in agriculture history and mode of action / J. J. Dibner, J. D. Richards // Poult Sci. – 2005. – Vol. 84. – Р. 634–643.
- Dibner, J. J. Use of organic acids as a model to study the impact of gut microflora on nutrition and metabolism / J. J. Dibner, P. Buttin // J Appl Poultry Res. – 2002. – Vol. 11. – P. 453–463.
- Dicks, L. M. T. Characterization and Cloning of the Genes Encoding Enterocin 1071A and Enterocin 1071B, Two Antimicrobial Peptides Produced by Enterococcus faecalis BFE 1071 / L. M. T. Dicks, E. Balla, M. Toit // Appl. Environ. Microbiol. – 2013. – Vol. 93, № 4. – P. 1298–1304.
- Diep, D. B. Characterization of the locus responsible for the bacteriocin production in Lactobacillus plantarum C11 / D. B. Diep, L. S. Havarstein, I. F. Nes // J. Bacteriol. – 1996. – Vol. 178. – P. 4472–4483.
- Divercin V41, a new bacteriocin with two disulphide bonds produced by Carnobacterium divergens V41: primary structure and genomic organization /
A. Métivier, M. Pilet, X. Dousset, O. Sorokine, P. Anglade, M. Zagorec, J. Piard,
D. Marion, Y. Cenatiempo, E. Fré maux // Microbiology. – 1998. – Vol. 144. –
P. 2837–2844. - Diversity and succession of the intestinal bacterial community of the maturing broiler chicken / J. Lu, U. Idris, B. Harmon, C. Hofacre, J. J. Maurer,
M. D. Lee // Appl Environ Microbiol. – 2003. – Vol. 69. – Р. 6816–6824. - Effect of antibiotic withdrawal in feed on chicken gut microbial dynamics, immunity, growth performance and prevalence of foodborne pathogens / S. Kumar [et al.] // PloS ONE. – 2018. – Vol. 13. – e0192450.
- Effect of bacillus subtilis cgmcc 1.1086 on the growth performance and intestinal microbiota of broilers / Y. Li, Q. Xu, Z. Huang, L. Lv, X. Liu, C. Yin,
H. Yan, J. Yuan // J. Appl. Microbiol. – 2016. – Vol. 120. – Р. 195–204. - Effect of buturic acid on perfomance, gastrointestinal tract helth and carcass characteristics in broiler chickens / A. K. Panda, S. V. Rama, M. N. Raju,
G. Shyam Sunder // Asian-Australasin Journal Animal Science. – 2009. – Vol. 22. – P. 1026–1031. - Effect of carbohydrase and protease on growth performance and gut health of young broilers fed diets containing rye, wheat, and feather meal / F. Yan,
J. J. Dibner, C. D. Knight, M. Vazquez-Anon // Poult Sci. – 2017. – Vol. 96(4). –
Р. 817–828. - Effect of citric acid supplemented diets on aflatoxin degradation, growth perfomance and serum parameters in broiler chickens / L. Salgado-Transito,
J. C. Rio-Garcia, J. L. Moreno-Martinez, A. Mendez-Albores // Archivos de Medicina Veterinaria. – 2011. – Vol. 43. – P. 215–222. - Effect of combination of citric acid and microbial phytase on digestibility of calcium, phosphorous and mineralization parameters of tibia bone in broilers /
Y. E. Nezhad, J. G. Gale-Kandi, T. Farahvash, A. R. Yeganeh // African Journal Biotechnology. – 2011. – Vol.10. – P. 15089–15093. - Effect of dietary supplementation of prebiotics and probiotics on intestinal microarchitecture in broilers reared under cyclic heat stress / S. Ashraf, H. Zaneb,
M. S. Yousaf, A. Ijaz, M. U. Sohail, S. Muti, M. M. Usman, S. Ijaz, H. Rehman //
J. Anim. Physiol. Anim. Nutr. – 2013. – Vol. 97. – Р. 68–73. - Effect of formic acid on performance, digestibility, intestinal histomorphology and plasma metabolite levels of broiler chickens / F. Hernandez, J. Madrid, V. Garcia, J. Orendo, M. D. Megias // British Poultry Science. – 2006. – Vol. 47. –
P. 50–56. - Effect of supplementation of prebiotic mannan-oligosaccharides and probiotic mixture on growth perfomance of broilers subjected to chronic heat stress /
M. U. Sohail, M. E. Hume, J. A. Byrd, D. J. Nisbet, A. Ijaz, M. Z. Shabbir,
H. Rehman // Poultry Science. – 2012. – Vol. 91. – P. 2235–2240. - Effects of dietary fat source and subtherapeutic levels of antibiotic on the bacterial community in the ileum of broiler chickens at various ages / A. Knarreborg, M. A. Simon, R. M. Engberg, B. B. Jensen, G. W. Tannock // Appl Environ Microbiol. – 2002. – Vol. 68. – Р. 5918–5924.
- Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers / Z. R. Xu, C. H. Hu, M. S. Xia, X. A. Zhan, M. Q. Wang // Poult Sci. – 2003. – Vol. 82. – Р. 1030–1036.
- Effects of dietary incusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens /
W. A. Awad, K. Ghareeb, S. Abdel-Raheem, J. Bohm // Poultry Science. – 2009. – Vol. 88. – P. 49–55. - Effects of dietary prebiotics, probiotics and synbiotics on perfomance, caecal bacterial populations and caecal fermentation concentrations of broiler chickens / S. Mookiah, C. C. Sieo, K. Ramasamy, N. Abdullah, Y. W. Ho // J Science Food Agriculture. – 2014. – Vol. 94. – P. 341–348.
- Effects of enterococcus faecium and dried whey on broiler performance, gut histomorphology and intestinal microbiota / H. E. Samli, N. Senkoylu, F. Koc,
M. Kanter, A. Agma // Arch. Anim. Nutr. – 2007. – Vol. 61. – Р. 42–49. - Effects of galacto-oligosaccharides and a Bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens /
S. J. Jung, R. Houde, B. Baurhoo, X. Zhao, B. H. Lee // Poult Sci. – 2008. –
Vol. 87. – Р. 1694–1699. - Effects of swine gut antimicrobial peptides on the intestinal mucosal immunity in specific pathogen –free chickens / D. Wang, W. Ma, R. She, Q. Sun,
Y. Liu, Y. Hu, L. Liu, Y. Yang, K. Peng // Poultry Science. – 2009. – Vol. 88. –
P. 967–974. - Effects of synbiotic feed additive in comparison to antibiotic growth promoter on perfomance and health status of broilers / M. Mohnl, Y. Acosta Aragon,
A. Acosta Ojeda, B. Rodriguez Sanchez, S. Pasteiner // Poultry Science. – 2007. – Vol. 86 (suppl. 1). – P. 217. - Error-correcting barcoded primers for pyrosequencing hundreds of samples in multiplex / M. Hamady, J. J. Walker, J. K. Harris, N. J. Gold, R. Knight // Nat Methods. – 2008. – Vol. 5. – Р. 235–237.
- European Parliament. EC 248/97. Regulation of the European Parliament and of the Council concerning novel foods and novel food ingredients : Official Journal. – 1997. – Vol. 14. – Р. 1–7.
- Farias, R. N. Letters in Purification and N-terminal acid sequence of enterocin CRL35, a pediocin-like bacteriocin produced by Enterococcus faecium CRL35 / R. N. Farias, M. E. Farias, R. Holgado // Appl. Microbiol. – 2005. – Vol. 56. –
P. 417–419. - Feed-additive probiotics accelerate yet antibiotics delay intestinal microbiota maturation in broiler chicken / P. Gao, C. Ma, Z. Sun, L. Wang, S. Huang,
X. Su, J. Xu, H. Zhang // Microbiome. – 2017. – Vol. 5. – Р. 91. - Fimland, G. Comparative studies of immunity proteins of pediocin-like bacteriocins / G. Fimland, Eijsink, J. Nissen’Meyer // Microbiology. – 2002. –
Vol. 148. – P. 3661–3670. - Fosgerau, K. Peptide therapeutics: current status and future directiuons /
K. Fosgerau, T. Hoffmann // Drug Discovery Today. – 2015. – Vol. 20. –
P. 122–128. - Franz, C. M. Production and characterization of enterocin 900, a bacteriocin produced by Enterococcus faecium BFE 900 from black olives / C. M. Franz,
U. Schillinger, W. H. Holzapfel // Int. J. Food Microbiol. – 1996. – Vol. 29. –
Р. 255–270. - Gaggia, F. Probiotics and prebiotics in animal feeding for safe food production / F. Gaggia, P. Mattarelli, B. Biavati // Inter J Food Microbiol. – 2010. –
Vol. 141. – Р. 15–28. - Goh, H. F. Purification and characterization of bacteriocin produced by Weissella confusa A3 of Dairy Origin / H. F. Goh, K. Philip // PLoS One. – 2015. – Vol. 10.
- Gulluce, M. Bacteriocins: promising antimicrobials. Microbial pathogens and strategies for combating them / M. Gulluce, M. Karadayi, O. Baris // In: Science, Technology and Education / A. Mendes-Vilas (Ed.). – FORMATEX, Madrid, Spain, 2013. – Р. 1016–1027.
- Hecht, H. Structural characterization of sodium alginate and calcium
alginate / H. Hecht, S. Srebnik // Biomacromolecules. ‒ 2016. ‒ Vol. 17, № 6. ‒ P. 2160‒2167. - Hegde, N. V. Comparison of antimicrobial resistant genes in chicken gut microbiome grown on organic and conventional diet / N. V. Hegde, S. Kariyawasam, C. DebRoy // Veter Anim Sci. – 2016. – Vol. 1. – Р. 9–16.
- Henderson, J. Purification and primary structure of pediocin PA-1 produced by Pediococcus acidilactici PAC-1.0 / J. Henderson, A. Chopko, P. Dick van Wassenaar // Arch. Biochem. Biophys. – 1992. – Vol. 295. – P. 5–12.
- Heterologous expression of the bacteriocin mesentericin Y105 using the dedicated transport system and the general secretion pathway / F. Biet, J. M. Berjeaud, R. W. Worobo, Y. Cenatiempo, C. Fremaux // Mircobiology. – 1998. –
Vol. 144. – Р. 2845–2854. - Host and Environmental factors affecting the intestinal microbiota
in chickens / J. G. Kers, F. C. Velkers, E. A. J. Fischer, G. D. A. Hermes,
J. A. Stegeman, H. Smidt // Front Microbiol. – 2018. – Vol. 9. – Р. 235. - Human nutrition, the gut microbiome and the immune system / A. L. Kau, P. P. Ahern, N. W. Griffin, A. L. Goodman, J. I. Gordon // Nature. – 2011. –
Vol. 474. – Р. 327–336. - Hurst, A. Nisin / A. Hurst // Adv. Appl. Microbiol. – 1981. – Vol. 27. –
P. 85–123. - Huyghebaert, G. An update on alternatives to antimicrobial growth promoters for broilers / G. Huyghebaert, R. Ducatelle F. Van Immerseel // Veterinary
J. – 2011. – Vol. 187. – P. 182–188. - Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials / V. A. Torok, R. J. Hughes, L. L. Mikkelsen, R. Perez-Maldonado, K. Balding, R. MacAlpine, N. J. Percy,
K. Ophel-Keller // Appl. Environ. Microbiol. – 2011. – Vol. 77. – Р. 5868–5878. - Immune response of chicken gut to natural colonization by gut microflora and to Salmonella enterica serovar enteritidis infection / M. Crhanova [et al.] // Infect Immun. – 2011. – Vol. 79. – Р. 2755–2763.
- Influence of Salmonella enterica serovar enteritidis infection on the composition of chicken cecal microbiota / P. Videnska, F. Sisak, H. Havlickova, M. Faldynova, I. Rychlik // BMC Veter Res. – 2013. – Vol. 9. – Р. 140.
- Insights into broilers’ gut microbiota fed with phosphorus, calcium and phytase supplemented diets / D. Borda-Molina, M. Vital, V. Sommerfeld, M. Rodehutscord, A. Camarinha-Silva // Front Microbiol. – 2016. – Vol. 7. – Р. 2033.
- Insights into the biodiversity of the gut microbiota of broiler chickens /
L. Mancabelli [et al.] // Environ Microbiol. – 2016. – Vol. 18 – Р. 4727–4738. - Intestinal function and gut microflora of broiler chickens as influenced by cereal grains and microbial enzyme supplementation / M. D. Shakouri, P. A. Iji,
L. L. Mikkelsen, A. J. Cowieson // J Anim Physiol Anim Nutrit. – 2009. – Vol. 93. – Р. 647–658. - Jiang, H. Heterologous expression and purification of plantaricin NC8,
a two-peptide bacteriocin against Salmonella spp. from Lactobacillus plantarum ZJ316 / H. Jiang, P. Li, Q. Gu // Protein Expr. Purif. – 2016. – Vol. 127. – Р. 28–34. - Juturu, V. Heterologous protein expression in Pichia pastoris: Latest research progress and applications / V. Juturu, J. C. Wu // Chembiochem. – 2018. – Vol. 19. – Р. 7–21.
- Kim, J.W. Dietary organic acids for broiler chickens: a review / J. W. Kim, J. H. Kim, D. Y. Kil // Colombian Journal Animal Science and Veterinary Medicine. – 2015. – Vol. 28. – P. 109–123.
- Kogut, M. H. Spatial and temporal changes in the broiler chicken cecal and fecal microbiomes and correlations of bacterial taxa with cytokine gene expression / M. H. Kogut, B. B. Oakley // Front Veter Sci. – 2016. – Vol. 3. – Р. 11.
- Kohl, K. D. Diversity and function of the avian gut microbiota /
K. D. Kohl // J. Compar Physiol B Biochem Syst Environ Physiol. – 2012. –
Vol. 182. – Р. 591–602. - Kraus, A. Molecular Mechanisms of Bacterial Resistance to Antimicrobial Peptides / A. Kraus, D. Peschel. – Berlin; Heidelberg, 2006. – P. 231–250.
- Kumar, S. Revolution in rumen microbiology / S. Kumar, D. W. Pitta //
In: Rumen Microbiology: From Evolution to Revolution / A. K. Puniya, R. Singh,
D. N. Kamra, ediotrs. – New Delhi : Springer, 2015. – Р. 357–379. - Kumar, V. Halocin HA1: an archaeocin produced by the haloarchaeon Haloferax larsenii HA1 / V. Kumar, S. K. Tiwari // Process Biochem. – 2017. –
Vol. 61. – Р. 202–208. - Lactobacillus alvi sp. nov., isolated from the intestinal tract of chicken /
H. J. Kim, S. J. Eom, S. J. Park, C. J. Cha, G. B. Kim // FEMS Microbiol Lett. – 2011. – Vol. 323. – Р. 83–87. - Lactococcin MMFII, a novel class IIa bacteriocin produced by Lactococcus lactis MMFII, isolated from a Tunisian dairy product / M. Ferchichi, J. Frère, K. Mabrouk, M. Manai // FEMS Microbiol. Lett. – 2001. – Vol. 205. – P. 49–55.
- Lagkouvardos, I. Cultured microbes represent a substantial fraction of the human and mouse gut microbiota / I. Lagkouvardos, J. Overmann, T. Clavel //
Gut Microb. – 2017. – Vol. 8. – Р. 493–503. - Lee, S. G. Purification and characterization of mejucin, a new bacteriocin produced by Bacillus subtilis SN7 / S. G. Lee, H. C. Chang // LWT Food. Sci. Technol. – 2018. – Vol. 87. – Р. 8–15.
- Lopez-Lara, I. M. Bacterial lipid diversity / I. M. Lopez-Lara, O. Geiger // Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2017. – Vol. 1862. – Р. 1287–1299.
- Maldonado, A. Purification and genetic characterization of plantaricin NC8, a novel coculture-inducible two-peptide bacteriocin from Lactobacillus plantarum NC8 / A. Maldonado, J. Ruiz’Barba, R. Jiménez’Díaz // Appl. Environ. Microbiol. – 2003. – Vol. 69. – P. 383–389.
- Markowiak, P. The role of probiotics, prebiotics and synbiotics in animal nutrition / P. Markowiak, K. Slizewska // Gut Pathog. – 2018. – Vol. 10. – Р. 21.
- Martinez, B. Lactococcin 972 : a homodimeric lactococcal bacteriocin whose primary target is not the plasma membrane / B. Martinez, J. E. Suarez, A. Rodriguez // Microbiology. – 1996. – Vol. 142. – P. 2393–2398.
- MiSeq: A Next Generation Sequencing Platform for Genomic Analysis /
R. K. Ravi, K. Walton, M. Khosroheidari // Methods Mol Biol. – 2018. – Vol. 1706. – Р. 223–232. – doi : 10.1007/978-1-4939-7471-9_12. Review. - Modifying the gastrointestinal ecology in alternatively raised poultry and the potential for molecular and metabolomic assessment / S. H. Park, I. Hanning,
A. Perrota, B. J. Bench, E. Alm, S. C. Ricke // Poult Sci. – 2013. – Vol. 92. –
Р. 546–561. - Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment / J. L. Danzeisen, H. B. Kim,
R. E. Isaacson, Z. J. Tu, T. J. Johnson // PLoS ONE. – 2011. – Vol. 6. – e27949. - Molecular analysis of bacterial populations in the ileum of broiler chickens and comparison with bacteria in the cecum / J. Gong [et al.] // FEMS Microbiol Ecol. – 2002. – Vol. 41. – Р. 171–179.
- Molecular approaches to the analysis of gastrointestinal microbial ecosystems / H. J. Flint [et al.] // In: Molecular Approaches to the Analysis of Gastrointestinal Microbial Ecosystems / G. C. Perry, editor. – Oxford : CABI Publishing, 2006. – Р. 107–123.
- Muriana, P. M. Purification and partial characterization of lactacin F, a bacteriocin produced by Lactobacillus acidophilus 11088 / P. M. Muriana, T. R. Klaenhammer // Appl. Environ. Microbiol. – 1991. – Vol. 57. – P. 114–121.
- Next generation sequencing: the future of molecular genetics in poultry production and food safety / S. Diaz-Sanchez, I. Hanning, S. Pendleton, D. D’souza // Poult Sci. – 2013. – Vol. 92. – Р. 562–572.
- Nilsen, T. Enterolysin A, a cell wall degrading bacteriocin from Enterococcus faecalis LMG 2333 / T. Nilsen, I. F. Nes, H. Holo // Appl. Environ. Microbiol. – 2003. – Vol. 69. – Р. 2975–2984.
- Nisin is an effective inhibitor of Clostridium difficile vegetative cells and spore germination / C. Le Lay, L. Dridi, M. G. Bergeron, M. Ouellette, I. Fliss //
J. Med. Microbiol. – 2016. – Vol. 65. – Р. 169–175. - Nuclear Magnetic Resonance Structure and Mutational Analysis of the Lactococcin A Immunity Protein / P. E. Kristiansen, C. Persson, V. Fuochi, A. Pedersen, G. B. Karlsson, J. Nissen-Meyer, C. Oppegård // Biochemistry. – 2016,
Nov 15. – Vol. 55(45). – Р. 6250–6257. - Nucleotide and amino acid sequence of pap-gene (pediocin AcH production) in Pediococcus acidilactici H. / A. Motlagh, A. Bhunia, F. Szostek, T. R. Hansen, M. C. Johnson, B. Ray // Lett. Appl. Microbiol. – 1992. – Vol. 15. – P. 45–48.
- Oscariz, J. C. Classification and mode of action of membrane-active bacteriocins produced by gram-positive bacteria / J. C. Oscariz, A. G. Pissabarro // Int. Microbiol. – 2011. – № 18. – P. 13–19.
- Pan, D. Intestinal microbiome of poultry and its interaction with host and diet / D. Pan, Z. Yu // Gut Microbes. – 2014. – Vol. 5. – Р. 108–119.
- Parameters and techniques to determine intestinal health of poultry as constituted by immunity, integrity, and functionality / S. H. Jeurissen, F. Lewis,
J. D. Van der Klis, Z. Mroz, J. M. Rebel, A. A. Ter Huurne // Curr Issues Intest Microbiol. – 2002. – Vol. 3. – Р. 1–14. - Patten, J. D. Use of organic acids in broiler diets / J. D. Patten, P. W. Waldroup // Poultry Sci. – 1988. – Vol. 67. – P. 1178–1182.
- Performance comparison of benchtop high-throughput sequencing platforms / N. J. Loman [et al.] // Nat Biotechnol. – 2012. – Vol. 30. – Р. 434–439.
- Pourabedin, M.; Guan, L.; Zhao, X. Xylo-oligosaccharides and virginiamycin differentially modulate gut microbial composition in chickens / M. Pourabedin,
L. Guan, X. Zhao // Microbiome. – 2015. – Vol. 3. – Р. 15. - Prevalence of temporomandibular disorder pain in Chinese adolescents compared to an age-matched Swedish population / L. Hongxing, A. N. Astrøm,
T. List, I. M. Nilsson, A. Johansson // J Oral Rehabil. – 2016, Apr. – Vol. 43(4). –
Р. 241–248. – doi : 10.1111/joor.12366. - Probiotics in Animal Nutrition: Production, Impact and Regulation /
Y. S. Bajagai, A. V. Klieve, P. J. Dart, W. L. Bryden. – FAO : Rome, Italy, 2016. - Production of pediocin PA–1 in the methylotrophic yeast Pichia pastoris reveals unexpected inhibition of its biological activity due to the presence of collagen–like material / L. Beaulieu, D. Groleau, C. B. Miguez, J. F. Jette, H. Aomari,
M. Subirade // Protein. Expr. Purifi. – 1999. – Vol. 43. – Р. 111–125. - Purification and amino acid sequence of lactocin 705, a bacteriocin produced by Lactobacillus casei CRL 705 / J. Palacios, G.Vignolo, M. E. Farias,
A. R. de Ruis Holgado, G. Oliver, F. Sesma // Microbiol. Res. – 1999. – Vol. 154. – P. 199–204. - Purification and characterization of plantaricin A, a Lactobacillus plantarum bacteriocin whose activity depends on the action of two peptides / J. Nissen-Meyer, A. G. Larsen, K. Sletten, M. Daeschel, I. F. Nes // J. Gen. Microbiol. –
1993. – Vol. 139. – P. 1973–1978. - Purification and cloning of sakacin 674, a bacteriocin from Lactobacillus sake Lb674 / A. Holck, L. Axelsson, K. Huhne, L. Krockel // FEMS Microbiol Lett. – 1994. – Vol. 115(2-3). – Р. 143–149.
- Purification and partial characterization of a novel bacteriocin synthesized by Lactobacillus paracasei HD1-7 isolated from Chinese sauerkraut juice / J. Ge,
Y. Sun, X. Xin, Y. Wang, W. Ping // Sci. Rep. – 2016. – Vol. 6. – 19366. - Purification, characterization and amino acid sequencing of divergicin M35: a novel class IIa bacteriocin produced by Carnobacterium divergens M35 /
I. Tahiri, M. Desbiens, R. Benech, E. Kheadr, C. Lacroix, S. Thibault, D. Ouellet,
I. Fliss // Int. J. Food Microbiol. – 2004. – Vol. 97. – P. 123–136. - QIIME allows analysis of high-throughput community sequencing data /
J. G. Caporaso [et al.] // Nat Methods. – 2010. – Vol. 7. – Р. 335–336. - Ranganath, B. S. Optimisation of media for the growth and production of bacteriocin from Bacillus coagulans / B. S. Ranganath, T. Sharmila, B. V. Balasubramanyam // Res. Technol. Sci. Engine. – 2015. – Vol. 1. – Р. 109–114.
- Rappe, M. S. The uncultured microbial majority / M. S. Rappe, S. J. Giovannoni // Ann Rev Microbiol. – 2003. – Vol. 57. – Р. 369–394.
- Response of broiler chicken to dietary supplementation of organic acids /
S. Adil, T. Banday, G. A. Bhat, M. Salahuddin, M. Raquib, S. Shanaz // J Central Eur Agriculture. – 2011. – Vol. 12. – P. 498–508. - Review: supplemention of phytase and carbohydrases to diet for poultry / T. A. Woyengo, C. M. Nyachoti // Canadian J Animal Science. – 2011. – Vol. 91. – P. 177–192.
- Sakacin g, a new type of antilisterial bacteriocin / L. Simon, C. Fremaux, Y. Cenatiempo, J. M. Berjeaud // Appl. Environ. Microbiol. – 2002. – Vol. 68. –
P. 6416–6420. - Sakacin-A antimicrobial packaging for decreasing Listeria contamination in thin-cut meat: preliminary assessment / A. Barbiroli, A. Musatti, G. Capretti,
S. Iametti, M. Rollini // J Sci Food Agric. – 2017. – Vol. 97(3). – Р. 1042–1047. - Samanta, S. Comparative efficacy of an organic acid blend and bacitracin methylene disalicylate as growth promoters in broiler chickens: effect on performance, gut histology, and small intestinal milieu / S. Samanta, S. Haldar,
T. K. Ghost // Vet Med International. – 2010. – 645150. - Scientific committee. introduction of a qualified presumption of safety (QPS) approach for assessment of selected microorganisms referred to EFSA1. Opinion of the Scientific Committee (Question No EFSA-Q-2005-293) // EFSA J. –
2007. – Vol. 587. – Р. 1–16. - Selective enrichment of bifidobacteria in the intestinal tract of broilers by thermally produced ketoses and effect on broiler performance / J. A. Patterson,
J. I. Orban, A. L. Sutton, G. N. Richards // Poult Sci. – 1997. – Vol. 76. – Р. 497–500. - Sharma, D. Simultaneous production of biosurfactants and bacteriocins by probiotic Lactobacillus casei MRTL3 / D. Sharma, B. Singh Saharan // Int. J. Microbiol. – 2014. – 698713 (7 pages).
- Simple and rapid purification of pediocin PA-1 from Pediococcus pentosaceous NCDC 273 suitable for industrial application / V. B. Simha, S. K. Sood,
R. Kumariya, A. K. Garsa // Microbiol. Res. – 2012. – Vol. 167. – Р. 544–549. - Siragusa, G. R. Brochocin-C, a new bacteriocin produced by Brochothrix campestris / G. R. Siragusa, C. N. Cutter // Appl. Environ. Microbiol. – 1993. –
Vol. 59. – Р. 2326–2328. - Streptomyces scopuliridis sp. nov., a bacteriocin-producing soil streptomycete / M. H. Farris, C. Duffy, R. H. Findlay, J. B. Olson // Int. J. Syst. Evol. Microbiol. – 2011. – Vol. 61. – Р. 2112–2116.
- Structural analysis and characterization of lacticin Q, a novel bacteriocin belonging to a new family of unmodified bacteriocins of gram-positive bacteria /
K. Fujita, S. Ichimasa, T. Zendo, S. Koga, F. Yoneyama, J. Nakayama, K. Sonomoto // Appl. Environ. Microbiol. – 2007. – Vol. 73. – P. 2871–2877. - Sultimova, T. D. Bacteriocins Of Lactic Acid Bacteria / T. D. Sultimova,
E. V. Zakharov // Newsletter of Essutm. – 2016. – Vol. 2(59). – P. 41–47. - Surugau, L. N. Peptide separation by capillary electrophoresis with ultraviolet detection: some simple approaches to enhance detection sensitivity and resolution / L. N. Surugau // The Malaysian Journal of Analytical Sciences. – 2011. –
Vol. 15(2). – P. 273–287. - Suzuki, K. New aspects of IgA synthesis in the gut / K. Suzuki, A. Nakajima // Inter Immunol. – 2014. – Vol. 26. – Р. 489–494.
- Tax4Fun: predicting functional profiles from metagenomic 16S rRNA data / K. P. Abhauer, B. Wemheuer, R. Daniel, P. Meinicke // Bioinformatics. – 2015. – Vol. 31. – Р. 2882–2884.
- The chicken gastrointestinal microbiome / B. B. Oakley [et al.] // FEMS Microbiol Lett. – 2014. – Vol. 360. – Р. 100–112.
- The continuing story of class IIa bacteriocins / D. Drider, G. Fimland,
Y. Hechard, L. M. McMullen, H. Prevost // Microbiol. Mol. Biol. Rev. – 2006. –
Vol. 70. – Р. 564–582. - The development of bactericidal yeast strains by expressing the Pediococcus acidilactici pediocin gene (pedA) in Saccharomyces cesevisiae / H. Schoeman, M. A. Vivier, M. Dutoit, L. M. T. Dicks, I. S. Pretorius // Yeast. – 1999. – Vol. 15. – Р. 647–656.
- The effects of different thermal treatments and organic acid llevels in feed on microbial composition and activity in gastrointestinal tract of broilers / F. Boroojeni, W. Vahjen, A. Mader, F. Knorr, I. Ruhnke, I. Rohe, A. Hafees, C. Villodre, K. Manner, J. Zentek // Poultry Science. – 2014. – Vol. 93. – P. 1440–1452.
- The membrane topology of immunity proteins for the two-peptide bacteriocins carnobacteriocin XY, lactococcin G, and lactococcin MN shows structural diversity / A. P. Britton, S. R. van der Ende, M. J. van Belkum, L. A. Martin-Visscher // Microbiologyopen. – 2019. – e957. – doi : 10.1002/mbo3.957.
- The microbiome of the chicken gastrointestinal tract / C. J. Yeoman,
N. Chia, P. Jeraldo, M. Sipos, N. D. Goldenfeld, B. A. White // Anim Health Res Rev. – 2012. – Vol. 13. – Р. 89–99. - The potency of the broad spectrum bacteriocin, bactofencin A, against staphylococci is highly dependent on primary structure, N-terminal charge and disulphide formation / P. M. O’Connor, E. F. O’Shea, P. D. Cotter, C. Hill, R. P. Ross // Sci. Rep. – 2018. – Vol. 8. – Р. 11833.
- The two-peptide class II bacteriocins: structure, production, and mode of action / Oppegard, P. Rogne, L. Emanuelsen, P. E. Kristiansen, G. Fimland, J. Nissen-Meyer // J. Mol. Microbiol. Biotechnol. – 2007. – Vol. 13. – Р. 210–219.
- The two-peptide class II bacteriocins: structure, production, and mode of action / C. Oppegard, P. Rogne, L. Emanuelsen, P. E. Kristiansen, G. Fimland,
J. NissenMeyer // J. Mol. Microbiol. Biotechnol. – 2007. – Vol. 13. – Р. 210–219. - Thermophilin 13, a nontypical antilisterial poration complex bacteriocin, that functions without a receptor / O. Marciset, M. C. Jeronimus’Stratingh, B. Mollet, B. Poolman // J. Biol. Chem. – 1997. – Vol. 272. – P. 14277–14284.
- Thompson, J. K. Characterization of a proteinaceous antimicrobial produced by Lactobacillus helveticus CNRZ450 / J. K. Thompson, M. A. Collins,
W. D. Mercer // J. Appl. Bacteriol. – 1996. – Vol. 80. – Р. 338–348. - Tiehaczek, P. S. Cloning and sequencing of curA encoding curvacin A, the bacteriocin produced by Lactobacillus curvatus LTH1174 / P. S. Tiehaczek,
R. F. Vogel, W. P. Hammes // Arch. Microbiol. – 1993. – Vol. 160. – P. 279–283. - Updating benchtop sequencing performance comparison / S. Junemann
[et al.] // Nat Biotechnol. – 2013. – Vol. 31. – Р. 294–296. - US patent. US20100129864, A1. Novel bacteriocins, transport and vector system and method of use thereof / E. Stiles Michael, C. Vederas John, J. van Belkum Marius, W. Worobo Randy, J. Worobo Rodney, K. McCormick John,
G. Greer Gordon, M. McMullen Lynn, J. Leisner Jorgen, Alison Poon, Charles
Franz. – MAP, 2010. - US patent. US20100286030 A1. Novel bacteriocin from a new streptomyces species / M. H. Farris, P. F. Churchill, J. B. Olson. – 2010.
- US Patent. US20140148379 A1. Control of unwanted bacteria in fermentation systems with bacteriocin / M. Liu, E. J. Summer. – 2014.
- US Patent. US20150320829 A1. Broad spectrum bacteriocin for control of unwanted Bacteria / M. Liu, E. J. Summer. – 2015.
- US patent. US6403082 B1. Bacteriocins, transport and vector system and method of use thereof / E. Stiles Michael, C. Vederas John, J. van Belkum Marius, W. Worobo Randy, J. Worobo Rodney, K. McCormick John, G. Greer Gordon,
M. McMullen Lynn, J. Leisner Jorgen, Poon Alison, Franz Charles. – MAP, 2002. - US patent. US7960505 B2. Lantibiotics and uses thereof / D. J. O’Sullivan, J. H. Lee. – 2011.
- US patent. US9326523 B2. Class I and II lantibiotics from geobacillus thermodenitrificans / W. A. Van der Donk, N. Garg, Y. Goto, W. Tang. – 2016.
- Van Reenen, C. Isolation, purification and partial characterization of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum / C. Van Reenen, L. Dicks, M. Chikindas // J. Appl. Microbiol. – 1998. – Vol. 84. – P. 1131–1137.
- Variations of phosphorous accessibility causing changes in microbiome functions in the gastrointestinal tract of chickens / B. Tilocca, M. Witzig, M. Rodehutscord, J. Seifert // PLoS ONE. – 2016. – Vol. 11. – e0164735.
- Vijayakumar, P. P. A Microplate Growth Inhibition Assay for Screening Bacteriocins against Listeria monocytogenes to Differentiate Their Mode-of-Action / P. P. Vijayakumar, P. M. Muriana // Biomolecules. – 2015. – Vol. 5(2). – Р. 1178–1194.
- Waite, D. W. Exploring the avian gut microbiota: current trends and future directions / D. W. Waite, M. Taylor // Front Microbiol. – 2015. – Vol. 6. – Р. 673.
- Wassels, S. The lactic acid bacteria, the food chain, and their regulation / S. Wessels, L. Axelsson, E. B. Hansen // Trends in Food Science and Technology. 2004. № 15. P. 498505.
- Wei, S. Bacterial census of poultry intestinal microbiome / S. Wei,
M. Morrison, Z. Yu // Poult Sci Sympos. – 2013. – Vol. 92. – Р. 671–683. - Willey, M. Lantibiotics: peptides of diverse structure and function /
M. Willey, W. van der Donk // Ann. Rev. Microbiol. – 2007. – Vol. 61. –
P. 477–501. - Yanagida, F. Isolation and characterization of lactic acid bacteria from soils in vineyards / F. Yanagida, Y. Chen, T. J. Shinohara // Gen. Appl. Microbiol. – 2005. – Vol. 52. – P. 21–28.
- Yang, Y. Dietary modulation of gut microflora in broiler chickens: a review of the role of six kinds of alternatives to in-feed antibiotics / Y. Yang, P. A. Iji, M. Choct // World Poult Sci J. – 2009. – Vol. 65. – Р. 97.
- Yegani, M. Factors affecting intestinal health in poultry / M. Yegani,
D. R. Korver // Poult Sci. – 2008. – Vol. 87. – Р. 2052–2063. - Zendo, T. Lactococcal membrane-permeabilizing antimicrobial peptides / T. Zendo, F. Yoneyama, K. Sonomoto // Appl. Microbiol. Biotech. – 2010. –
Vol. 88. – P. 1–9. - Zhao, W. Antibiotic resistance genes in China: occurrence, risk, and correlation among different parameters / W. Zhao, B. Wang, G. Yu // Environ Sci Pollut Res Int. – 2018. – Vol. 25(22). – Р. 21467–21482.
- Zhou, W. Functional cloning and characterization of antibiotic resistance genes from the chicken gut microbiome / W. Zhou, Y. Wang, J. Lin // Appl Environ Microbiol. – 2012. – Vol. 78. – Р. 3028–3032.