
Титульный лист и исполнители
РЕФЕРАТ
Отчет: 71 стр., 1 кн., 7 рис., 24 табл., 131 источник литературы.
КРУПНЫЙ РОГАТЫЙ СКОТ, КОСТРОМСКАЯ ПОРОДА, ОТБОР, ПОДБОР, ЭКСТЕРЬЕР, ПРОДУКТИВНОЕ ДОЛГОЛЕТИЕ, ГЕНЕТИЧЕСКИЙ ПОТЕНЦИАЛ, ИММУНОГЕНЕТИЧЕСКИЕ МАРКЕРЫ, ПЦР, КАППА-КАЗЕИН, БЕТА-КАЗЕИН, АДГЕЗИЯ ЛИМФОЦИТОВ.
Объектом исследования являлся крупный рогатый скот костромской породы в племенных хозяйствах Костромской области.
Цель работы – сохранение и улучшение генофонда костромской породы крупного рогатого скота по племенным и продуктивным качествам с использованием различных методов селекции.
В процессе работы был проведен анализ генофонда костромской породы по племенным и продуктивным качествам с использованием традиционных и молекулярно-генетических методов селекции.
По результатам работы с линиями и родственными группами получены новые данные. Впервые начата работа по экстерьерному профилированию быков-производителей костромской породы. Проведенный генетический мониторинг в стадах костромской породы показал, что за период с 1977 по 2018 гг. число аллелей сократилось с 42 до 20. Это обстоятельство вызывает необходимость разработки специальной селекционной программы, обеспечивающей сохранение в стаде необходимого генетического разнообразия с целью сохранения оптимального уровня гетерозиготности. Впервые проведен анализ полиморфизма гена бета-казеина в популяции скота костромской породы, который показал высокое распространение желательного генотипа А2А2.
В ходе научно-исследовательской работы создана чувствительная и быстрая тест-система ПЦР в реальном времени на определение мутации BLAD в племенных хозяйствах области. Система прошла апробацию на маточном поголовье в племенных стадах по разведению скота костромской, ярославской, черно-пестрой (улучшенной голштинской) пород. В стадах по разведению черно-пестрого скота 6,0% маточного поголовья являются носителями мутации BLAD.
Степень внедрения – племенным хозяйствам по разведению скота костромской породы даны рекомендации по использованию различных методов селекции. В племенных хозяйствах по разведению черно-пестрого скота животные-носители мутации BLAD поставлены на контроль.
ТЕРМИНЫ И ОПРЕДЕЛЕНИЯ
В настоящем отчете о научно-исследовательской работе применены следующие термины с соответствующими определениями:
Аллель — одна из возможных структурных состояний гена. Аллели гена находятся в парных хромосомах (одна из которых наследуется от матери, другая — от отца) и определяют направление развития одного и того же признака.
Антиген — вещество, которое воспринимается организмом как чужеродное и вызывает специфический иммунный ответ.
Ген — наследуемая часть генома, оказывающая влияние на какой-либо фенотипический признак.
Гетерозиготность — наличие в организме одной или нескольких пар различающихся аллелей.
Гетерогенность групп животных (генотипическая) —неоднородность составляющих групп особей, обусловленная разнообразием генотипов в пределах одной группы.
Гомозиготность — наличие в генотипе организма одной или нескольких пар одинаковых аллелей.
Гомогенность групп животных генотипическая — однородность составляющих групп особей, обусловленная однообразием генотипов в пределах группы.
Генетический контроль (мониторинг) — контроль вредных мутаций, включающий тщательный клинический анализ болезней и уродств, экспертизу происхождения аномальных животных, выяснение роли наследственности в их этиологии.
Дезоксирибонуклеиновая кислота — генетический материал, который определяет наследственные характеристики большинства живых организмов.
ДНК-маркеры — участки нуклеиновой последовательности ДНК, отличающиеся полиморфизмом и тесно связанные с генами, отвечающими за нужный признак.
Мутация (лат. mutatio — изменение) — стойкое, то есть такое, которое может быть унаследовано потомками, изменение генома.
Полиморфизм — гены, которые представлены в популяции несколькими разновидностями аллелей, что обуславливает разнообразие признаков внутри вида.
ПЕРЕЧЕНЬ СОКРАЩЕНИЙ И ОБОЗНАЧЕНИЙ
В настоящем отчете о научно-исследовательской работе применены следующие сокращения и обозначения:
ДНК — дезоксирибонуклеиновая кислота (deoxyribonucleic acid, DNA)
ДТТ — дитиотреитол (dithiothreitol, DTT)
КРС — крупный рогатый скот
ПДРФ — полиморфизм длин рестрикционных фрагментов (Restriction Fragment Length)
П.Н. — пара нуклеотидов (base pair, b.p.)
ПЦР — полимеразная цепная реакция (Polymerase Chain Reaction, PCR)
ЭДТА — этилендиаминтетрауксусная кислота (ethylenediaminetetraacetic acid, EDTA)
CSN3 — ген каппа-казеина
CSN2 — ген бета-казеина
BLAD — синдром дефицита адгезии лимфоцитов
ВY — брахиспинальный синдром
ПХИ — количество (дней) лет от начала лактации до выбытия животного из стада
РИБ — родительский индекс быка
РИК — родительский индекс коровы
РИД — родительский индекс дочери
ВВЕДЕНИЕ
Вопрос о необходимости сохранения генофонда сельскохозяйственных животных впервые в нашей стране был поднят еще в 20-х годах. Уже тогда наиболее дальновидные ученые и практики оценили перспективы его использования для создания заводских пород и типов животных, а также для селекции будущего. Этому способствовало то обстоятельство, что многие местные отродья обладали рядом уникальных свойств и могли проявлять их в таких условиях кормления и содержания, в которых культурные западные породы просто не выживали, отмечают Сервах Б.А., Рахматулина Н.Р. [87].
С начала 90-х годов XX столетия в молочном скотоводстве России идут два четко выраженных процесса: сокращение поголовья и расширение использования мирового генофонда (главным образом, голштинской и бурой швицкой пород). Результатом этих процессов явилось сокращение численности отечественных пород, пишет Кузнецов В.М. [48].
Костромская порода крупного рогатого скота – одна из лучших отечественных пород. Она прошла сложный путь создания и совершенствования, началу которого способствовали экономические (близость крупных рынков сбыта) и природные условия (богатые сенокосами поймы рек). Костромская порода адаптирована к местным климатическим условиям, имеет крепкую конституцию, продуктивное долголетие. Молоко костромских коров является ценным технологическим сырьем для получения сыров. Порода отличается устойчивостью к таким заболеваниям, как туберкулез, бруцеллез и лейкоз, хорошо адаптирована к условиям интенсивной технологии, сообщает Всяких А.С. [16].
В 2018 году поголовье крупного рогатого скота на территории области составило 9317 голов, в том числе 5841 голов костромской породы, или 0,35% от всего поголовья крупного рогатого скота, разводимого на территории нашей страны. В Костромской области сосредоточено основное поголовье скота костромской породы (64,70%) с продуктивностью по первой лактации – 5718,00 кг молока с содержанием жира 4,10% и 3,21% белка, по третьей и старше – 6003,00 кг молока, 4,07% жира и 3,21% белка. В современных условиях отечественным породам трудно конкурировать с обильно-молочными коммерческими породами.
Именно акцент на качественные показатели молока, долголетие, устойчивость к ряду заболеваний и поддержка со стороны государства дает шанс на сохранение и рациональное использование малочисленных отечественных пород.
1 ОБЗОР ЛИТЕРАТУРЫ
1.1 Характеристика генеалогических корней бурого скота
В молочном скотоводстве страны около 5,0% породного скота относится к бурым породам. Бурый скот неприхотлив к условиям содержания и разведения, а высокая адаптационная способность к различным природно-климатическим зонам, наряду с крепкой конституцией и устойчивостью к заболеваниям, способствует его распространению и использованию для получения молока и мяса высокого качества [131].
Современные бурые породы крупного рогатого скота являются производными бурой швицкой породы. Все они имеют бурую масть и генетическое сходство со швицами. Бурые породы разводят в 74 странах на всех континентах нашей планеты. Такое внимание к этому скоту обусловлено не только его высокими акклиматизационными способностями, но и хорошими молочными и откормочными качествами, устойчивостью к ряду заболеваний, Дунин И.М., Харитонов С.Н., Ермилов А.Н., Сперанский А.Т. и др. [28].
При его создании основной целью было получить животных мясо-молочного направления продуктивности с хорошими рабочими качествами. Позднее (XIX век), в связи с ростом спроса на молоко и молочные продукты, были выведены молочные типы и породы бурого скота.
В Российской Федерации на основе скрещивания с бурыми производителями были выведены костромская и кавказская бурая породы. До середины 70-х годов отечественные бурые породы России совершенствовались по самостоятельным планам и программам, которыми предусматривалось продолжение селекции животных в направлении улучшения как мясных, так и молочных качеств. Однако с 1975 года начинается активное использование швицких быков из США с целью ускорения темпов генетического улучшения молочных качеств отечественного бурого скота. Этот период, видимо, следует считать началом преобразования отечественных бурых пород комбинированного направления продуктивности в специализированный молочный скот.
Швицкий бурый скот США произошел от бурого скота Швейцарии. Первая партия швицких животных была завезена в США в 1869 г. Кларком, Скоттом и Харрисом в штат Массачусетс. С этого времени бурый скот начинает довольно активно размножаться и распространяться.
К середине шестидесятых годов прошлого столетия его численность превысила 500 тыс. голов. Многие авторы, исследовавшие бурых швицев США, отмечают выдающуюся роль коровы по кличке Джейн оф Верном 28496 в становлении современной популяции этой породы, Дунин И.М., Харитонов С.Н., Ермилов А.Н., Сперанский А.Т. и др. [28].
Генеалогическая структура производителей бурого американского скота приведена на рисунке 1.
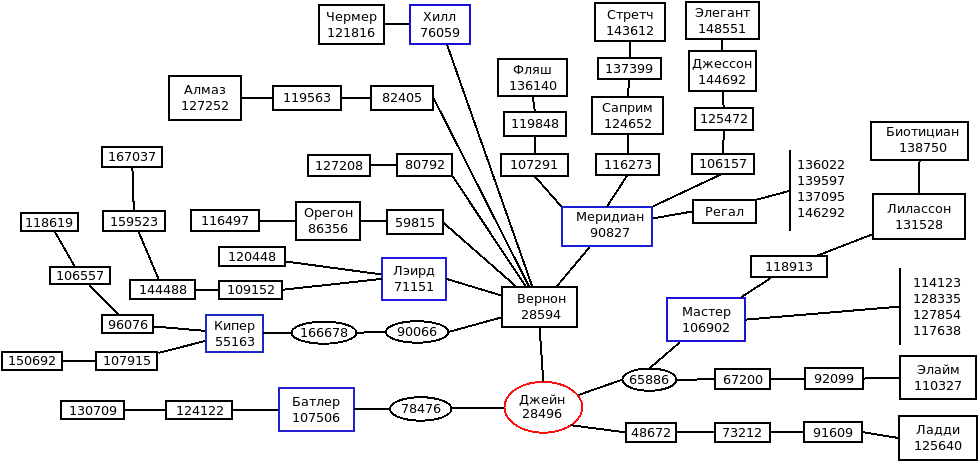
Рисунок 1 – Генеалогическая структура производителей бурого американского скота (И.М. Дунин, 1998)
Она родилась в 1928 году на ферме Орбека штата Висконсин. Эксперты того времени признавали, что Джейн оф Верон обладала лучшим экстерьером и особенно выменем. В швицкой породе культивируется 37 линий, но наибольшее распространение получили группы Мастера 106902, Хилла 76059, Меридиана 90827, Концентрата 106157, Лэирда 71151 [28].
1.2 Выведение и этапы создания скота костромской породы
Костромская порода крупного рогатого скота прошла сложный путь создания и совершенствования, началу которого способствовали экономические (близость крупных рынков сбыта) и природные условия (богатые сенокосами поймы рек), пишет Е.Г. Емельянов [29].
По мнению Ружевского А.Б., Рубана Ю.Д. и Бердника П.П. [82], порода формировалась в семь этапов.
На первом этапе (начало ХIХ века) местный скот скрещивали с завозимыми быками холмогорской породы. В результате этой работы было создано костромское отродье холмогорского скота.
На втором этапе (вторая половина ХIХ века) в Костромскую губернию завозили быков-производителей айрширской, вильстермаршской, симментальской породы, швицкого и альгауского отродья бурого швицкого скота. В результате были сформированы две помесные группы – мисковский и бабаевский скот. Мисковский скот формировался в верховьях реки Костромы (зона маслоделия) под влиянием айрширской и вильстермашской пород. По описанию П.Г. Власова, этот тип молочно-мясного скота был представлен животными красно-пестрой (72,0%), красной (13,5%), белой (11,5%), буро-пестрой (1,4%) и черно-пестрой масти (1,0%).
Средняя живая масса коровы составляла 380,00-390,00 кг. Бабаевский скот (назван по Николо-Бабаевскому монастырю) образовывался при активном использовании альгауского отродья и незначительном – швицких производителей. Коровы одноцветной серебристо-желтой масти бабаевского скота были распространены в селениях по течению реки Солоницы в Тетеринской волости Нерехтского уезда и в Шунгенской волости Костромского уезда, Горский Н.А. [22].
Третий этап (с 1899 по 1910 гг.) характеризуется возрастающим масштабом использования быков швицкой породы, завозимых из Швейцарии и стада Петровско-Разумовской сельскохозяйственной академии, в результате чего создается значительный массив помесных животных, Шаумян В.А. [111].
В течение четвертого этапа (1911—1912 гг.) повторно завозятся швицкие быки и используются для поглотительного скрещивания с помесями первого поколения Ружевский А.Б., Рубан Ю.Д., Бердник П.П. [82].
Пятый этап (1912—1918 гг.) характеризуется продолжением активного использования швицких производителей, главным образом на бабаевской группе животных.
В течение шестого этапа (1918—1929 гг.) в ряде хозяйств организуются племенные фермы. В 1920 году был организован совхоз «Караваево», в котором начали вести углубленную племенную работу по созданию высокопродуктивного стада. В 1925—1926 гг. в хозяйство были завезены швицкие быки из стада Московской сельскохозяйственной академии им. К.А. Тимирязева, в том числе бык Артист ИШ-55, с которым связана генеалогическая «революция» в породе, по данным Шалугина Б.В. [109, 113].
Седьмой этап (1930—1940 гг.) знаменателен тем, что начинается работа по созданию новой породы. В 1932 году совхоз «Караваево» преобразовали в племенное хозяйство, провели оценку племенных качеств животных, организовали учет происхождения и продуктивности. В 1934 году был организован Костромской госплемрассадник швицкого скота на базе племенных ферм Костромского и Нерехтского районов.
В 1944 году комиссия Министерства сельского хозяйства СССР провела апробацию поголовья в племенном заводе «Караваево» и Костромском госплемрассаднике. После тщательного изучения документов и осмотра животных в натуре комиссия приняла решение об апробации новой породы крупного рогатого скота, присвоив ей название «костромская». Авторами породы признаны С.И. Штейман, В.А. Шаумян, Н.А. Горский, А.Д. Митропольская, П.А. Малинина. После утверждения породы начинается интенсивное распространение костромского скота. По данным А.С. Всяких [16], во второй половине 40-х годов XX столетия породу начинают разводить в Ивановской, Владимирской, Витебской, Могилёвской областях, Татарской и Марийской АССР. Костромских животных завозят в Тульскую и Смоленскую области, где скрещивают со швицким скотом, и в Киргизскую ССР, где используют при выведении алатауской породы.
В 1970—1980 гг. в костромской породе проводилось формирование новых заводских линий. Родоначальниками линий становятся Курс ИКС-161 и Модный КТКС-630 (внуки Каро КТКС-101). В Ивановской и Владимирской областях образуется родственная группа Клеверка ИКС-196 (сын Хлора КТКС-200) и родственная группа Спутника ВДКС-134 (внук Хлора КТКС-200).
В 1980—2013 гг. в породе формируются пять родственных групп на основе бурых швицких быков американской селекции и маточного поголовья костромской породы с использованием производителей родственных групп – Батлера 107506, Концентрата 106157, Мастера 106902, Меридиана 90827, Хилла 76059, отмечает Емельянов Е.Г. [29].
В настоящее время среди проверяемых быков наибольший удельный вес (73,5%) приходится на долю родственных групп, сформированных на основе швицкой породы. Быки линий костромской породы составили всего лишь 26,5%, по данным Гусевой Т.Ю. [26].
Таким образом, отечественные породы – это не только стратегический фонд для развития сельского хозяйства, но и неотъемлемая часть нашего культурного наследия.
1.3 Традиционные методы селекции молочного скота
1.3.1 Отбор и подбор в молочном скотоводстве
В практике племенной работы отбор и подбор теснейшим образом связаны, это последовательные звенья единого процесса, направленного на непрестанное качественное совершенствование отдельных стад и целых пород животных в избранном направлении. Отбор решает судьбу особи: будет или не будет она участвовать через свое потомство в дальнейшей эволюции породы, а подбор определяет качество будущего потомства.
Большое внимание подбору уделяли в своих научных работах и практике известные ученые: Щепкин М.М. [114], Кулешов П.Н. [50, 15], Богданов Е.А. [10], Кисловский Д.А. [40], Кравченко Н.А. [44, 45], Гребень Л.К. [23], Лось Н.Ф. [66], Марченко Г.Г. [70] и др.
Однако причины неодинаковой сочетаемости различных заводских линий, маточных семейств и отдельных животных в пределах линии или породы до сих пор остаются нераскрытыми, Борисенко Е.Я. [11]. Но первая гипотеза не объясняет всего многообразия различной сочетаемости при подборе. Гипотеза сверхдоминирования – т.е. такого взаимодействия, когда гетерозигота лучше любой исходной гомозиготы. По этой гипотезе гетерозигота будет лучше гомозигот в тех случаях, когда доминантный и рецессивные гены выполняют различные функции. Новый вариант гипотезы сверхдоминирования – гипотеза «физиологического равновесия», т.е. взаимодействия между генными продуктами (ростовыми веществами) [7]. По мнению многих генетиков, перечисленные выше гипотезы охватывают основные формы взаимодействия наследственных факторов и могут объяснить различную сочетаемость при подборе, а также явление гетерозиса.
В начале XIX в. были определены два основных типа подбора: однородный и разнородный. Суть однородного подбора заключается в подборе животных со схожими продуктивными качествами. Его цель –сохранить у потомков высокие племенные качества родителей [9]. Крайним вариантом однородного подбора является инбридинг. Без всякого преувеличения можно сказать, что проблема инбридинга интересует в настоящее время широкий круг специалистов – биологов, генетиков, зоотехников-селекционеров, которые понимают все сложности, связанные с изучением генетической природы инбридинга, с противоречивыми порой результатами их использования в практике животноводства, Ивлев Ф.В. [32], Москаленко Л., Коновалов А. [72].
Биологическая сущность инбридинга и его практический смысл, писал Кулешов П.Н. [51], сводится к повышению наследственной устойчивости (препотентности) инбредного потомства, или, проще говоря, к «сгущению крови» лучших родителей [119]. Примером удачного использования инбридинга в селекции в указанный период времени могут служить успехи английских заводчиков при создании новых пород: Р. Бэквел – создатель лейстерской породы овец; братья Р. и Ч. Коллинги – шортгорнской мясной породы крупного рогатого скота, Тулей – крупной белой породы свиней и др. Близкородственное спаривание применялось при создании новой американской породы санта-гертруда, которая ведет свое происхождение от одного быка – Манки, Ивлев Ф.В. [32].
Значение инбридинга особенно возрастает в условиях крупномасштабной селекции при получении быков-производителей и создании их линий. При этом опасность в этих случаях уменьшается, так как все быки, включая инбридированных, проверяются по качеству потомства –это непременное условие крупномасштабной селекции, отмечают Эрнст Л.К., Зиновьева Н.А. [115].
Например, в своей работе Parland S.M., Kearney J.F., Rath M., Berry D.P. [124] определи влияние инбридинга на производство молока, число соматических клеток, плодовитость, выживаемость, показатели отела у ирландских коров голштино-фризской породы. Они отмечают, что инбридинг имел пагубное влияние на молочную продуктивность коров, показатели отела, плодовитость и выживаемость молодняка.
Любимов А.И. с соавторами [67, 68] провели сравнительный анализ продуктивных и воспроизводительных качеств аутбредного и инбредного скота, разводимого в племенных заводах Удмуртской Республики и Свердловской области. Три четверти случаев инбридинга в Свердловской области приходится на долю отдаленного и одна четверть – на умеренный. На близкий и тесный инбридинг приходится не более 1 %, это может говорить о крайне осторожном отношении селекционеров Свердловской области к близким степеням инбридинга.
Донник И.М., Мымрин В.С., Лоретц О.Г. и др. [27] приводят данные, что в Свердловской области от разведения коров с умеренной степенью инбридинга в племенных хозяйствах недополучено 150 т молока в год.
Лягин Ф.Ф. [67] отмечал, что из 1011 коров-рекордисток по костромской породе стада ГПЗ «Караваево» 443 получены при различных степенях родственного спаривания: из них 36 голов (8,1%) – при тесном родстве, 93 головы (21,0%) –при близком, 198 голов (44,7%) – при умеренном и 116 голов (26,2%) – при отдаленном родстве. Оценка близкородственного разведения крупного рогатого скота костромской породы показала, что с целью консолидации определенных положительных качеств лучше применять умеренный и отдаленный инбридинг и только в исключительных случаях, для получения быков-производителей линий, прибегать к близким степеням родства, Федосенко Е.Г., Баранов А.В., Тараканова Г.Н. и др. [99].
Суть гетерогенного (разнородного) подбора состоит в том, что спариваемые животные заведомо различаются по признакам подбора.
Цель гетерогенного подбора – увеличение изменчивости признаков, повышение продуктивности, резистентности, репродуктивных качеств, поглощение недостатков, имеющихся у отдельных животных или групп, Борисенко Е.Я. [11].
Анализ количественных и качественных показателей коров голштинской породы в ПЗ «Повадино» Московской области показал, что среди всех кроссов линий лучшим по количеству молока следует считать кросс Силинг Трайджун Рокит 252803 × Монтвик Чифтейн 95679.
При внутрилинейном подборе лучший удой 8266,00 кг молока был отмечен у потомков линии Силинг Трайджун Рокит, однако это на 269,00 кг меньше, чем у коров кросса Силинг Трайджун Рокит 252803 × Монтвик Чифтейн 95679 [94].
Для совершенствования ряда пород по молочной продуктивности (бестужевская, холмогорская, ярославская, истобенская и симментальская) применяют скрещивание с голштинской породой. При этом имеются животные, полученные как с прилитием крови (50,0%), так и с высокой долей крови (50,0% и более) по голштинской породе. Фактически не осталось племенных стад, где можно было бы получить чистопородных холмогорских быков от высокопродуктивных коров [110].
Зоотехническая наука разработала общий подход к обоснованию племенного подбора, но пока еще не позволяет безошибочно предвидеть результаты каждого спаривания.
Таким образом, в настоящее время зоотехнику-селекционеру приходится на основе анализа результатов оценки производителей по качеству потомства, предыдущего подбора и племенной работы в целом сначала осторожно и с некоторым риском, а потом все более обоснованно намечать наиболее обещающие сочетания отдельных групп и особей и планировать дальнейшую племенную работу.
1.3.2 Значение в племенной работе экстерьерной оценки коров
Зоотехническая наука уделяет особое внимание изучению связи продуктивных качеств сельскохозяйственных животных с их конституциональными и экстерьерными особенностями. Абрампальский Ф.Н. [2] считает, что желая добиться быстрого повышения молочной продуктивности у отечественного скота, селекционеры обращают основное внимание на удой, совершенно забывая при этом об экстерьере животного, а между тем форма и функция домашних животных нераздельны.
Внешний вид и продуктивность представляют единое целое и являются выражением обмена веществ. Известно, что для образования одного литра молока необходимо, чтобы через вымя коровы прошло 400-500 литров крови [3, 17, 18].
Рахматулина Н.Р., Егиазарян A.B., Сервах Б.А. [78] отмечают, что уделяя большое внимание при селекции молочной продуктивности, ленинградские селекционеры в последние годы придают большое значение организации линейной экстерьерной оценке коров в племенных стадах и экстерьерному профилированию быков-производителей. Корова молочного направления продуктивности имеет своеобразное строение.
Тип телосложения коров, ориентированный на выносливость и высокую продуктивность, играет важную роль для эффективного производства продукции.
Попов Н.В. [75], Салбырин Р. [83], Сарапкин В.Г., Бялькина Т.А. [84], Стрекозов Н., Сельцов В., Кожухов Д. [90] отмечают, что при селекции только на молочную продуктивность ухудшается переднее прикрепление вымени, его глубина увеличивается, что приводит к сокращению использования коров в стаде. Поэтому необходима комплексная оценка быков-производителей, с учётом продуктивных и экстерьерных качеств дочерей. Это позволит повысить эффективность молочного скотоводства за счёт длительного использования наиболее высокопродуктивных животных. Своевременное выявление и исключение из селекционного процесса быков с серьёзными недостатками и пороками экстерьера их дочерей позволяет улучшить молочный тип животных, отмечают Бургомистрова О.Н. [14], Ефимова Л. [30], Завертяев Б.П. [31], Шишкина Н.В. [112], Ржепаковский В.В., Баранник Г.В. [81].
По мнению авторов Гринь М.П., Якусевич А.М., Буткевич С.К. и др. [24], Карликова Д.В., Щеглова Е.В., Казарбина Д.Р. и др. [38], Новикова A.B. [73], Рахматулиной Н.Р. с соавторами [76, 77, 79, 80], оценка быков по качеству потомства в одинаковых условиях кормления и содержания путём одновременного сравнения между собой позволяет выявить производителя- улучшателя как молочной продуктивности, так и воспроизводительных качеств маточного поголовья.
Сотрудники лаборатории оценки генотипа ГНУ ВНИИГРЖ Россельхозакадемии Логинов Ж.Г. с соавторами [57, 58, 61], Рахматулина Н.Р., Егиазарян А.В., Сервах Б.А. [78], Сервах Б.А., Рахматулина Н.Р. [87] разработали модели продуктивно-экстерьерных индексов для комплексной оценки быков-производителей. В 2009 году, при участии директора ВНИИГРЖ, изданы методические рекомендации комплексной оценки животных в молочном скотоводстве.
Логинов Ж.Г. с соавторами [59, 60, 65] указывают на то, что основные требования, предъявляемые классификатором к молочной корове при оценке, сводятся к следующему: молочная корова должна иметь лёгкую, сухую, слегка удлинённую голову; длинную и не толстую шею; глубокую удлинённую грудь; объёмное, сильно развитое, но не отвислое брюхо; ровный и хорошо развитый крестец; большое и хорошо прикреплённое к брюшной стенке вымя, оно должно быть достаточно развитым, но не отвислым, чашеобразной или ваннообразной формы с равномерно расположенными сосками одинакового размера, средними по величине и диаметру. Дно вымени должно находиться на 5-7 см выше скакательного сустава. Для молочной коровы характерно так называемое «открытое ребро». Кость ребра широкая и плоская. Рёбра широко посажены и наклонены от позвоночника вниз и назад, а между двумя последними рёбрами свободно помещаются 2-3 пальца. Бедро тощее плоское и широкое. Большое значение при оценке уделяется развитию и постановке конечностей и копыт.
Для определения производственного типа коров молочных и комбинированных пород Айсанов З. [4] предлагает учитывать условный объём туловища коровы, который рассчитывается исходя из ширины груди за лопатками, ширины в тазобедренных сочленениях, глубины груди и косой длины туловища.
Легошин Г.П. с соавторами [54] сообщают, что в отечественной и зарубежной науке и практике накоплено достаточно данных, свидетельствующих о связях оценок экстерьера и отдельных его статей с уровнем продуктивности и продолжительностью использования коров. Показано, что форма и функция вымени коров положительно коррелируют с удоями (r = 0,3 – 0,5), а разница в продуктивности коров крайних классов, например, черно-пестрой породы, по комплексной оценке телосложения (между «превосходно» и «удовлетворительно») превышает 2500,00 кг, или 40,0%. Ещё более впечатляет информация из Англии. Все первотёлки страны, получившие наивысшую комплексную оценку типа телосложения («очень хорошо»), имели средний удой за лактацию 8600,00 кг, а с оценкой «хорошо с плюсом» (на один класс ниже) — 7700,00 кг, или на 900,00 кг меньше.
Проблема увеличения сроков использования сельскохозяйственных животных всегда привлекала внимание селекционеров. Этому важному вопросу большое значение придавали основоположники советской зоотехнической науки: Лебедько Е.Я. [19], Непомнящий Г.А. [53], Лискун Е.Ф. [55], Логинов Ж.Г. с соавторами [56, 63, 64] и многие другие учёные. В таблице 1 приведены генетические корреляции между линейными экстерьерными признаками и продолжительностью хозяйственного использования животных (ПХИ), а также с удоем за 305 дней лактации [86, 93, 120].
Таблица 1 – Связь линейных экстерьерных признаков с удоем и продолжительностью хозяйственного использования (Short T.H., 1991)
| Наименование линейного признака экстерьера | Генетическая корреляция | |
| удой, за 305 дней лактации | ПХИ | |
| Рост | +0,01 | +0,06 |
| Крепость сложения | –0,04 | –0,11 |
| Глубина туловища | +0,07 | –0,09 |
| Молочные формы | +0,52 | +0,23 |
| Угол крестца | +0,13 | +0,04 |
| Ширина крестца | +0,02 | –0,03 |
| Задние ноги, вид сбоку | +0,06 | –0,02 |
| Прикрепление передней части вымени | –0,23 | +0,42 |
| Высота прикрепления задней части вымени |
+0,06 | +0,33 |
| Ширина задней части вымени | +0,16 | +0,28 |
| Борозда вымени | –0,01 | +0,34 |
| Глубина вымени | –0,41 | +0,44 |
| Размещение сосков | –0,03 | +0,28 |
| Финальная оценка | +0,06 | +0,41 |
Как видно из таблицы, наибольшую положительную связь с удоем имеют такие признаки экстерьера, как «молочные формы» и «ширина задней части вымени». В то же время такие признаки экстерьера, как «прикрепление передней части вымени» и «глубина вымени» связаны с удоем достоверной отрицательной корреляцией. Большинство линейных экстерьерных признаков связаны положительной достоверной связью с продолжительностью хозяйственного использования. К ним относятся: «молочные формы», «прикрепление передней части вымени», «высота прикрепления задней части вымени», «ширина задней части вымени», «борозда вымени», «глубина вымени», «размещение сосков».
По данным американских исследователей, корреляция между оценкой экстерьерного типа коров и продолжительностью их хозяйственного использования составляет от +0,41 до +0,54 [14, 112]. Связи между полученным доходом и балльной оценкой признаков приведены в таблице 2.
Таблица 2 – Корреляция между доходом от коровы и оценкой признаков типа (Cassele B.G., Pearson R.E., Stoeland J., Huemstra S., 1998 г.)
| Коррелирующие признаки | Племенное хозяйство | Товарное стадо |
| Доход – пожизненная продуктивность | 0,77* | 0,69* |
| Доход – рост | – | –0,21* |
| Доход – крепость | –0,19 | 0,24* |
| Доход – глубина туловища | –0,10 | –0,20* |
| Доход – молочный тип | 0,61* | 0,37* |
| Доход – наклон крестца | 0,24* | 0,09* |
| Доход – ширина крестца | 0,21* | 0,19* |
| Доход – расположение сосков | 0,27* | 0,15* |
Примечание: * – Р < 0,05.
В племенных стадах при высоком уровне продуктивности и физиологической нагрузке на организм большое значение имеют наклон крестца и расположение сосков. В стадах разной продуктивности самая тесная корреляция установлена между доходом и оценкой молочного типа в баллах.
Следовательно, чем выше оценка за молочный тип, тем выше доход. Это логично, так как выраженность молочного типа связана с затратами кормов на продукцию — чем четче выраженный молочный тип, тем большая часть энергии корма идет на производство молока.
Учет признаков типа делает более эффективной селекцию по продолжительности хозяйственного использования позволяет снизить частоту мастита, повышает производительность труда при доении коров.
1.3.3 Селекция молочного скота на продуктивное долголетие
Современное молочное животноводство характеризуется поступательным ростом продуктивности коров, который зачастую сопровождается сокращением срока хозяйственного использования. Это одна из значимых проблем в современном молочном скотоводстве – повышение срока производственного использования животных и получение от них высокой пожизненной продуктивности. Даже в лучших хозяйствах страны большинство животных не доживают до полной реализации своего потенциала молочной продуктивности [98]. Актуальность проблемы вызвана тем, что в большинстве регионов России сроки использования коров не превышают 3-3,5 лактаций, в то время как наивысшую продуктивность они обычно проявляют на 4-6 лактациях. Долголетие высокопродуктивных коров позволяет также ускорить селекционное улучшение стада [74].
Основные затраты в отрасли можно разделить на затраты по выращиванию ремонтного молодняка и затраты на содержание коровы. Причем окупаемость этих затрат неодинакова. Реализация молока в значительной части России рентабельна, в то время как реализация говядины от выбракованных коров и телок повсеместно убыточна. Рентабельность отрасли молочного животноводства зависит от того, на сколько прибыль от реализации молока превышает убытки от реализации выбракованных коров и телок [5, 92].
С увеличением продолжительности хозяйственного использования коров растет рентабельность. Так, по исследованиям Яранцевой С.Я. [116] выбывшие после первой лактации коровы обеспечивали рентабельность в 4,3%, при использовании коров в течение пяти лактаций рентабельность повышалась до 47,6%. Чем выше продолжительность хозяйственного использования коров, тем ниже издержки на выращивание ремонтных телок. Чем короче сроки использования коров, тем большее количество молодняка требуется для замены выбывающего поголовья. Так, в структуре стада должно быть на каждые 100 коров при ежегодном вводе 25,0% первотелок 74 телки и нетели, при вводе 30,0% первотелок – 87, при вводе 35,0% первотелок –108 [33].
То есть для увеличения экономических показателей следует стремиться к повышению удоя, как за лактацию, так и за весь период продуктивного использования. Конечно, в решении проблем продолжительности хозяйственного использования молочных коров и увеличения их пожизненной продуктивности огромную роль играют вопросы выращивания ремонтного молодняка. При оптимальном возрасте и массе тела при осеменении и первом отеле удается сократить выбраковку из-за низкой продуктивности [62, 92].
В России при оценке быков по качеству потомства процент потерь коров-первотелок практически не учитывается. В США и ряде стран Европы в оценку быка включаются все его дочери, имевшие продолжительность лактации более 40 дней. Там же при оценке быка по продолжительности хозяйственного использования включают всех его дочерей с продолжительностью лактации 15 дней и более, отмечает Кузнецов В.М. [48, 49].
Возможна также косвенная селекция по продолжительности хозяйственного использования, и она дает положительный результат. Жизнь коровы сокращают тяжелые отелы, влекущие за собой послеродовые заболевания, и как следствие – бесплодие. Много коров выбывает из-за болезней вымени и ног.
Еще 1988 г. [122] проанализирована связь оценки линейного типа коров по первой лактации с их продуктивностью и продолжительностью жизни. Были выявлены основные факторы, влияющие на эти признаки. Показано, что признаками, существенно и достоверно влияющими на молочную продуктивность, являются высота в крестце, выраженность молочного типа, а также высота и ширина вымени сзади. Положительно связаны с ПХИ слегка наклонный назад крестец, плотно прикрепленное спереди вымя, четко выраженная связка вымени, глубина вымени (положение дна вымени), вертикально поставленные соски.
В Германии были оценены по молочной продуктивности и показателям типа дочерей 2919 быков 1995—1997 гг. Позднее эти быки оценены по продолжительности использования их дочерей. Молочная продуктивность положительно коррелировала с молочным типом (r = +0,20) и с высотой задних долей (r = +0,15), отрицательно с глубиной вымени (r = –0,25).
Продолжительность использования коров находилась в положительной корреляции с оценкой глубины вымени (r = +0,30), оценкой скакательного сустава (r = +0,28), плотностью прикрепления передних долей (r = +0,25), положением – наклоном таза (r = +0,10). В то же время коровы ярко выраженного молочного типа имели более короткие сроки использования (r = –0,18), как и коровы с более глубоким туловищем (r = +0,20).
В молоке коров с удлиненными сосками (7 баллов и выше) было большое количество соматических клеток. Мертворожденные телята реже появляются у коров со слегка наклонным тазом [92].
Основное влияние на изменчивость продолжительности использования оказывают: оценка прикрепления вымени спереди, глубины вымени, постановка ног сбоку. Влияние признаков вымени на продолжительность использования было большим при беспривязном содержании и доении на доильных площадках.
Таким образом, высокопродуктивная корова с оптимальной продолжительностью использования должна иметь четко выраженный молочный тип, глубокое и широкое сзади, плотно прикрепленное спереди с четко выраженной бороздой средней глубины вымя с вертикально расположенными сосками, широкий и слегка наклонный крестец, правильную постановку ног и копыт.
1.4 Молекулярно-генетические методы в селекции молочного скота
1.4.1 Иммуногенетические маркеры
В нашей стране иммуногенетические исследования начаты с 1962 года, когда были созданы лаборатории по исследованию групп крови животных в Москве (ВИЖ), Ленинграде (ВНИИРГЖ), Новосибирске (СО АН СССР). Активно изучением групп крови крупного рогатого скота стали заниматься в ВИЖе с 1965 года после поездки П.Ф. Сорокового в лабораторию иммуногенетики Института физиологии и генетики чехословацкой академии наук. Сороковой П.Ф., Кузьминов И.П. [71, 88] проверили по результатам иммуногенетического анализа соответствия записей о происхождении животных и определили неправильную регистрацию данных у четверти изученных животных.
ФГУП «Московское» по племенной работе с 2001 года проводит иммуногенетический контроль происхождения крупного рогатого скота. Накопленный опыт работы лаборатории иммуногенетики ФГУП «Московское» по племенной работе показывает, что при сочетании традиционных зоотехнических и иммуногенетических методов удается свести до минимума ошибки в родословных, повысить технологическую дисциплину и уровень племенной работы, пишет Букаров Н.Г. с соавторами [12, 13].
Кольцов Д.Н., Дмитриева В.И. [42] провели мониторинг аллелей ЕАВ-локуса групп крови сычевской и голштинской пород красно-пестрой масти в хозяйствах Смоленской области
Стормонт К. [89], сравнивая по ЕАВ-системе группы крови пяти американских пород молочного скота, выявил, что наибольшую частоту имели аллели: OA’ (0,22), BO1Y2D’ (0,14), O1 (0,11), O1Y1E’3G’Y’ (0,10), O3QJ’K’O’ (0,04), у голштино-фризского скота: GY2E1 (0,20), OхQI’K’O’ (0,06), O1E’3 (0,06), у гернзейского скота: IOхE’1 (0,20), BGKOхF’O’ (0,13), у джерзейского скота: BGKOхY1A1F’3G’K’B’O’Y’ (0,24), O1T1Y2E’3F’K’ (0,18), OхE’3F’O’ (0,14), у швицкого скота: O1T1Y2E’3F’ (0,37), BO1Y2A’E’G’Y’ (0,10).
Для коров красной степной породы наиболее типичными являются аллели G2Y2Е’1Q», В1О2Y2D’, «b»и Q’. В геноме коров красной эстонской породы наиболее часто встречались аллели G2Y2E’tQ’ и B2G2. У коров красной горбатовской породы чаще других встречались аллели Y1Y» и «b», Князева Т.А. [41].
Под руководством Сахаутдинова И., Муратова Л., Исламова С. и др. [85] проводилась аттестация крупного рогатого скота симментальской породы Республики Башкортостан и изучения аллелофонда их групп крови. Часть животных была местной, часть – австрийской селекции.
Так, животные местной селекции имели аллели: O1G, I2QO’, Y1A’1Y’, Y1A1‘A’2Y’, Y1A’1A’2O’. Животные австрийской селекции аллели: Q’, E’3Q’, E’3Y’, Q’Y’ и E’3I’Q’. В стаде животных местной селекции ОПХ «Баймакское» обильномолочность коров обусловлена аллелями Y1A’1Y’, Y1A1‘A’2Y’, Y1A’1A’2O’, а высокая жирномолочность связана с присутствием аллеля I2QO’. Данные аллели могут быть использованы в селекционной работе в качестве генетических маркеров для раннего прогнозирования продуктивности животных. Среди коров австрийской селекции в данном хозяйстве желательны животные носители аллелей Q’, Е’3Q’, Е’3Y, Q’Y’ и Е’3I’Q’, а селекционный процесс необходимо направить на уменьшение численности животных с аллелями В2I2 и В2O’G2 [85].
Валитов Х.З., Карамаев С.В. [15] при анализе бестужевской породы СПК «Черновский» Смоленской области отмечают, что среди голштинизированных животных пожизненный удой выше среднего по стаду имели носительницы аллелей Y2G’Y’G», В2О1, B2I1P’2, P2Q, B2Y2G’G», P’2Q’E’2I’, B2G2KO’, G2Y2E’2Q’, B2G2KY2A’2О’, G2O1. Больше молока по сравнению со средним удоем по стаду в расчете на один день жизни было получено от носительниц аллелей В2О1, B2I1P2, P2QE’2I’, P’2Q’, B2G2KO’, G2Y2E’2Q’, B2G2KY2A’2О’, G2O1.
У коров симментальской породы, разводимых на территории Республики Башкортостан, концентрация аллелей А2‘; Y’; A2«Y’ и Р1А2«Y’ четко свидетельствовала о существовании положительной связи с обильномолочностью. Высокая жирномолочность коров данной популяции ассоциировала с аллелями А2«Е2‘,I’, Р1Е2‘,I’ и Р1Е2‘, Гумеров У., Исламова С. [25].
Ткаченко И.В., Гридин В.Ф. [96] установили, что в стадах крупного рогатого скота уральского типа иммуногенетическими маркерами высокого удоя являются аллели О1А’1, В2О1В’, Q’ и E’3F’2G’O’G», а наличие аллелей Е’1, G» и O3J’2К’O’ в генотипе животных на данном этапе селекционно-племенной работы связано с пониженной молочной продуктивностью.
Беляев В.И. с соавторами [8] изучали генетические маркеры устойчивости крупного рогатого скота к маститу. Авторами установлено, что частоты аллелей A, J, L, М, Z, С и S у восприимчивых и устойчивых к маститу коров примерно одинаковы. Далее авторы отмечают, что частота аллеля «b» достоверно выше у устойчивых коров, в то время как аллеля O1I’Q’ — у предрасположенных.
Одной из древних пород в Европе является швицкая бурая порода скота, выведенная в Швейцарии, а затем распространившаяся во многие страны мира. Опыт разведения бурой швицкой породы показывает, что это одна из наиболее лабильных в биологическом отношении порода крупного рогатого скота [100, 102, 104].
В Российском Нечерноземье эффективно разводятся швицкая и костромская породы бурого скота. Отечественный швицкий скот является результатом чистопородного разведения завезенных по импорту животных и поглотительного скрещивания местного скота с быками из Австралии, Венгрии, США и Швейцарии, отмечают Цысь В.И. с соавторами [101, 103, 105, 106], Лебедько Е.Я. [52], Кольцов Д.Н., Татуева О.В., Чернушенко В.К. и др. [43], Чернушенко В.К., Камошенков А., Бабичева В. [108], Красильникова Л.Н. [46], Абдулина Р.С., Гизатуллин Р.С., Салихов А.Р. [1].
При выведении «смоленского» типа бурого швицкого скота использовали быков американской селекции и частично джерсейских. Это привело к тому, что у «смоленских швицев» проявились ранее не встречавшиеся ЕАВ-аллели: B1O3Y2A’2E’3G’P’Q’Y’, B2G3QT1A’1P’, G1E3‘O’, G1OxO’, I1Y2E1‘GY’G», O1Q’, OxO’. Высокой продуктивностью отличались первотелки, в генотипе которых был аллель B1O3Y2A’2E’3G’P’Q’Y’, присущий швицкой породе американской селекции [39, 107].
Интенсивное использование швицких быков и их потомков способствовало изменению генетической структуры костромской породы. С момента использования быков швицкой породы в начале 70-х годов до настоящего времени произошло снижение частот аллелей Q, B2G3QT1A’1P’, I1G’G», B2G2KE’1F’2O’, P2I’, полностью элиминировали аллели О’, О1, Y2G’Y’G», B1G2KA’2B’O’, B2P2T2P’B», более широкое распространение получили аллели I1O1QA1‘, A1G’G», отмечает Глущенко М.А. [20].
Следовательно, аллельные гены групп крови отражают генетическую характеристику, пород, стад, линий, семейств и могут быть использованы при их совершенствовании достоверности происхождения племенных животных, контролем над изменением аллелофонда, выявления оптимальных параметров генетической изменчивости, создания групп животных, устойчивых к заболеваниям.
1.4.2 Полиморфизм генов молочных белков
В последнее время все большее значение в оценке генома животных приобретают молекулярно-генетические методы, входящие в систему биотехнологии. Созданы методы, которые позволяют изучать наследственность на уровне ДНК, Калашникова Л.А., Дунин И.М. [35, 36], Сулимова Г.Е. [91].
Работы по изучению полиморфных белков дали положительные результаты, однако выявленные корреляции с хозяйственно-полезными признаками в основном являются спецификой отдельных стад и пород животных. Эффективность анализа ограничивается только экспрессирующимися генами, например, по молочным белкам только у половозрастных лактирующих коров и невозможностью оценки различий в некодирующих областях генов, в том числе и в регуляторных областях. Эти сложности побуждают исследователей вести поиск новых систем генетического маркирования.
В этом направлении наиболее перспективны исследования непосредственно на уровне ДНК генома животных, в частности, по полиморфизму длин рестриктных фрагментов (ПДРФ). Задача поисков маркеров на уровне ДНК значительно упростилась с появлением метода амплификации ДНК с помощью полимеразной цепной реакции (ПЦР).
Казеины (от лат. caseus – сыр) составляют 80,0% от всех белков молока и относятся к наиболее ценным пищевым белкам, в состав которых входит полный набор незаменимых аминокислот. Кроме того, казеины являются источником пищевого кальция и фосфора, Кугенев П.В. и Барабанщиков Н.В. [47].
У крупного рогатого скота известно четыре основных типа казеинов — αS1, αS2, β и κ, Fox P. and Mullvichil D.M. [121]. В молоке они присутствуют в форме кальций-фосфатных мицелл (тончайших нитей), в стабилизации которых решающую роль играет каппа-казеин (κ-казеин). Это единственный белок, на который действует сычужный фермент, Holt C. and Sawyer L. [122].
Наиболее часто у КРС встречаются А- и В-аллельные варианты CSN3, различающиеся двумя аминокислотными заменами в 136 (Thr→Ile) и 148 (Asp→Ala) положениях полипептидной цепи, вызванными соответствующими точковыми мутациями в позициях 5309 (С→Т) и 5345 (A→ Тестирование) А- и В-аллелей представляет интерес с точки зрения практики, так как молоко коров, несущих в генотипе В-аллель каппа-казеина, имеет по сравнению с А-аллельным вариантом лучшую температурную устойчивость, более короткое время коагуляции, лучшую свертываемость и содержит мицеллы меньшего диаметра, что гораздо предпочтительнее для производства сыров [97, 117, 118, 123]. Они отмечают, что в молоке коров, несущих ВВ-генотип каппа-казеина, более высокое процентное содержание белка по сравнению с АА-генотипом каппа-казеина.
При производстве сыра из молока коров с генотипом ВВ каппа-казеина время коагуляции было короче на 24,0%, консистенция сгустка была лучше, а выход готовой продукции на 6,0% больше, чем при производстве сыра из молока коров, несущих гомозиготный генотип АА.
Схожие результаты наблюдались в работах Калашниковой Л.А. с соавторами [37], Бадина Г.А. и Шалугина Б.В. [6], Гончаренко Г.М. [21]. Европейская ассоциация животноводов и ряд крупных коммерческих фирм по животноводству предложили считать ВВ генотип каппа-казеина экономически важным селекционным критерием для пород КРС, специализированных в молочном направлении продуктивности [34].
По сообщению Тинаева А.Ш. [95] во многих странах мира, например, Германии, Дании, Голландии селекция по генотипам каппа-казеина включена в программы по разведению пород животных [121].
Ген бета-казеина имеет 12 аллельных вариантов, среди которых получили преимущества варианты А1 и А2. Они отличаются последовательностью аминокислот в первичной структуре: А1 содержит в 67-й позиции аминокислоту гистидин, тогда как вариант А2 – пролин.
Итак, различие в бета-казеинах было обнаружено в 1979 году. Но лишь в 1996 г. исследования показали, что эта разница важна. Работы в этой области вел новозеландский ученый Боб Элиот.
В то же самое время во Флориде врачи пришли к выводу о серьезности проблемы БКМ7 (бета-казоморфина-7), который оказывает вредное влияние на мозг человека. Они еще не знали про А1, но выяснили, что БКМ7 образуется при употреблении молока.
Результаты почти 6000 тестов на бета-казеин были предоставлены «Канадской молочной сети» (CDN). Примерная частотность генотипов по бета-казеину у разных пород представлена в таблице 3.
Таблица 3 – Примерная частотность генотипов по бета-казеину в разных породах животных
| Порода | Генотип | Количество тестированных животных | ||
| А2А2 | А1А2 | А1А1 | ||
| Голштинская | 35% | 49% | 16% | 4603 |
| Айрширская | 23% | 48% | 29% | 287 |
| Джерсейская | 65% | 32% | 3% | 752 |
| Бурая швицкая | 57% | 38% | 5% | 107 |
| Гернзейская | 53% | 19% | 28% | 145 |
В число пород с относительно высокой частотностью генотипа А2А2 входят джерсейская, бурая швицкая (бурые породы).
Селекция по маркерным генам каппа-казеина и бета-казеина позволит повысить белковомолочность, улучшить технологические качества молока, увеличить выход продуктов переработки, а также изменить качественный состав молока, которому пока что уделяется мало внимания.
1.4.3 Генетические аномалии и контроль за их распространением
Массовое использование глубокозамороженной спермы ограниченного числа производителей создает опасность распространения мутационных генов среди крупного рогатого скота. Производители, несущие в своем геноме дефекты, снижают воспроизводительные качества у потомков и в конечном итоге наносят большой экономический ущерб животноводству.
В отличие от доминантной мутации рецессивный мутационный ген может оставаться в популяциях в скрытом состоянии, не проявляя своего патологического действия. Только при скрещивании друг с другом двух особей, каждая из которых несет мутационный ген, могут выщепляться гомозиготы, которые и проявляют патологический эффект мутационного гена.
В международном списке летальных дефектов крупного рогатого скота содержится 402 наследственных фактора [127, 129]. Распространение дефектов по породам и странам разведения приведены в таблице 4.
Таблица 4 – Число генетических дефектов, обнаруженных в ряде пород крупного рогатого скота при разведении в разных странах
| Порода | Страна | Число генетических дефектов |
| Черно-пестрая | По всем странам | 31 |
| Германия | 3 | |
| Дания | 19 | |
| Венгрия | 1 | |
| Бельгия | 1 | |
| Польша | 3 | |
| Голштинская
(голштино-фризская) |
По всем странам | 78 |
| США | 30 | |
| Канада | 10 | |
| Германия | 7 | |
| Фризская | По всем странам | 51 |
| Айширская | По всем странам | 26 |
| Симментальская | По всем странам | 26 |
| Швицкая | По всем странам | 26 |
| США | 7 | |
| Европа | 14 |
Наследственные заболевания имеют разную степень распространенности в стадах крупного рогатого скота стран мира, но только целенаправленная работа по выявлению и выбраковке носителей мутационных генов позволяет эффективно бороться с их дальнейшим распространением [126, 125]. В результате анализа литературных и статистических данных выделены три генетические аномалии, наиболее часто встречаемые в отечественных популяциях крупного рогатого скота: CVM – комплексный порок позвоночника (до 20,0% по Эрнсту Л.К.), BLAD – синдром дефицита адгезии лимфоцитов (до 17,0% по Глазко В.И.) и ВY – брахиспинальный синдром (до 7,0% по данным ФГБУ «ВГНКИ»).
Серьезной проблемой, имеющей национальные масштабы, является дефицит адгезивности лейкоцитов крупного рогатого скота (BLAD). Животные, несущие в своем генотипе аллель в гомозиготном состоянии (CD18TL/TL), не способны противостоять вирусным и бактериальным инфекциям, что приводит к снижению иммунитета животных и заканчивается летальным исходом в первые месяцы развития.
Обычно взаимодействие клеток друг с другом, а также с экстрацеллюлярным матриксом на уровне молекул главным образом происходит через три семейства адгезивных белков: иммуноглобулинов, интегринов и селектинов. Семейство интегринов, в свою очередь, включает следующие факторы: LFA-1, Mac-1 и p150,95, известные в международной классификации как CD11/CD18 [145, 146].
Локус CD18 человека располагается на хромосоме 21q22.3, тогда как у крупного рогатого скота – на хромосоме 1 в группе синтении U10. У человека LAD является причиной нарушения адгезии в организме, аналогичная ситуация с синдромом гранулоцитопатии и у представителей крупного рогатого скота черно-пестрого генеалогического корня.
Kehrli M.-E.-Jr. et al. показали, что это связано с дефицитом Мас-1 гликопротеина [CD11b/CD18] лейкоцитов крови крупного рогатого скота и оно эквивалентно LAD человека, отсюда произошло ее название – BLAD.
Считается, что BLAD является уникальным случаем для сравнительной медицины человека и ветеринарной медицины при изучении LAD человека и других видов животных. Прекрасная модель для сравнительной генетики, по которой можно изучать клинику, фенотип и генетику самой болезни, – провести разработку диагностических подходов.
Это заболевание характеризуется резким снижением в лейкоцитах интегрина, что вызывает, в свою очередь, резкое снижение функциональной активности фагоцитоза, эндотелиальной адгезии, невозможности выхода лейкоцитов за пределы кровеносного сосуда и хемотаксиса. Лейкоциты больных BLAD содержат очень низкое количество поверхности, менее 2,0% от 160 кD белка здорового животного. Причиной заболевания является мутация CD18 гена, кодирующего гликопротеиновый интегрин [128, 130].
Более конкретно ситуация выглядит следующим образом: в данном локусе две мутации – одна «немая», в 775-м положении цитозин замещается на тимин, что ведет в 259-м положении аминокислотной последовательности замене лейцина на изолейцин. Другая мутация связана с заменой нуклеотида аденина на гуанин в 383-й позиции ДНК, вызывающая, в свою очередь, замещение в положении 128 аспарагиновой кислоты на глицин.
Течение болезни хроническое. Животные значительно отстают в росте и развитии, худеют при хорошем аппетите и очень восприимчивы к разного рода инфекциям. У телят часто наблюдается лишай. Одновременно отмечаются приступы лихорадки, постоянные нарушения в деятельности желудочно-кишечного тракта, а также признаки воспаления верхних дыхательных путей и легких. В большинстве случаев воспаляется область ротовой полости (гингивит). Попытка лечения приводит к исходным результатам, занимает много времени и в конечном итоге оказывается безуспешной.
Следовательно, для того чтобы не допустить дальнейшего бесконтрольного распространения мутации наряду с тестированием быков-производителей необходимо проводить тестирование коров быкопроизводящей группы и ремонтного молодняка. Введением системного мониторинга летальных мутаций возможно решить проблему повышения резистентности племенного поголовья и оздоровления селекционно-племенных стад.
2 ОСНОВНАЯ ЧАСТЬ
2.1 Материал и методика исследований
Объектом исследований являлись популяции молочного скота костромской породы, разводимой на территории Костромской области.
Материалом для лабораторных исследований послужили кровь коров костромской, ярославской, черно-пестрой (улучшенной голштинской) пород. Кровь отбирали из хвостовой вены в объёме 5 мл в вакуумные пробирки с сухим ЭДТА К3 (ООО «ГЕМ», Россия). Изучение аллелофонда по хозяйствам и породам по группам крови было проведено совместно с лабораторией иммуногенетики регионального информационно-селекционного центра.
Из цельной крови коров ДНК выделяли с использованием набора ПРОБА-ГС-Плюс (ОАО «ДНК-Технология», Россия), согласно методике, предоставленной фирмой-производителем. Принцип действия набора заключается в использовании лизирующего реагента, который предназначен для лизиса клеток, солюбилизации клеточного дебриса, а также для денатурации клеточных нуклеаз. Выделение ДНК проводили в изолированном боксе под ламинаром с использованием отдельного набора автоматических пипеток «Research» (Eppendorf, Германия) с целью избежания загрязнения образцов чужеродной ДНК. В ходе исследований разработана тест-система для быстрой и чувствительной идентификации однонуклеотидной замены (A→G) в позиции 383 гена CD18, ассоциированной с дефицитом адгезии лимфоцитов у коров (BLAD).
Определение генотипа основано на проведении полимеразной цепной реакции (ПЦР) с праймерами, общими для «дикого» и «мутантного» вариантов нуклеотидной последовательности. После завершения температурной программы ПЦР флуоресцентные зонды гибридизуются на полученную матрицу путем понижения температуры реакционной смеси. Для определения варианта последовательности использовались два типа олигонуклеотидов, гибридизующихся на матрицу рядом. Первый тип олигонуклеотидов помечен флуорофором, второй – гасителем флуоресценции.
Для генотипирования использовался один общий олигонуклеотид с гасителем флуоресценции и два сиквенс-специфичных олигонуклеотида, несущих различные флуорофоры (FAM-предковый аллель, VIC-мутантный аллель). Определение генотипа проводили путем измерения уровня флуоресценции в ходе температурной денатурации дуплексов олигонуклеотидов и полученных матриц. Результаты регистрировали в режиме реального времени. Если анализируемый образец содержит только один вариант нуклеотидной последовательности гена, т.е. гомозиготен по данному полиморфизму, температура плавления для зонда, образующего совершенный (полностью комплементарный) дуплекс, существенно выше, нежели для зонда, образующего несовершенный (частично некомплементарный) дуплекс.
При анализе гетерозиготного образца, содержащего оба варианта нуклеотидной последовательности, оба варианта зондов образуют совершенный дуплекс, поэтому температуры их плавления были практически одинаковы. Состав реакционной смеси приведен в таблице 5.
Таблица 5 – Компоненты реакционной смеси
| Компонент | Количество |
| ПЦР-буфер | 1-кратный |
| Смесь дезоксирибонуклеотидов | 25 мМ каждого |
| Зонд FAM | 0,1 пм/мкл |
| Зонд VIC | 0,1 пм/мкл |
| Зонд BHQ | 0,3 пм/мкл |
| Праймер 1 | 0,1 пм/мкл |
| Праймер 2 | 0,6 пм/мкл |
В реакции используется фермент Taq-полимераза в количестве 2,5 единицы активности на реакцию. Для предотвращения испарения амплификационной смеси на водную фазу реакции наслаивается 20 мкл минерального масла. Программа амплификации для детектирующих амплификаторов ДТ-48 и ДТ-96 (ООО «НПО ДНК-Технология») указана в таблице 6.
Таблица 6 – Программа амплификации для детектирующих амплификаторов ДТ-48 и ДТ-96
| №
блока |
Температура, °С | Мин. | Сек. | Число циклов | Режим оптических измерений | Тип блока |
| 1 | 80 | 0 | 30 | 1 | Цикл | |
| 94 | 1 | 30 | ||||
| 2 | 94 | 0 | 30 | 5 | Цикл | |
| 67 | 0 | 15 | V | |||
| 3 | 94 | 0 | 10 | 45 | Цикл | |
| 67 | 0 | 15 | V | |||
| 4 | 94 | 0 | 5 | 1 | Цикл | |
| 5 | 25 | 0 | 30 | 1 | Цикл | |
| 6 | 25 | 0 | 15 | 50 | V | Плавление |
| 7 | 10 | … | … | Хранение | Хранение |
Регистрация результатов ПЦР проводится автоматически программным обеспечением для детектирующего амплификатора ДТ-48 и/или ДТ-96. При возникновении недостоверных результатов необходимо повторить тест, начиная с процедуры выделения ДНК. Время проведения анализа – 80 минут. Тест-системы адаптированы к использованию с амплификаторами ДТ-48 и ДТ-96 (ЗАО «НПФ ДНК-Технология», Россия).
Специфичность тест-системы оценивали биоинформационным методом, сравнивая последовательность разработанных праймеров против базы данных геномных последовательностей Gen Bank и проверяя их на температурную стабильность и температуру отжига с помощью программного обеспечения Oligo 6.0. По результатам проверки ни один из праймеров не обладал достаточной степенью гомологии с геномом человека для того, чтобы привести к синтезу неспецифической мишени на расчётной температуре отжига. Чувствительность тест-системы оценивали методом серийных разведений. Чувствительность тест-системы составляет 50 копий ДНК на реакцию.
Тест-система демонстрирует стабильную воспроизводимость результатов при анализе количества копий ДНК, превышающего порог чувствительности тест-системы.
Оборудование и материалы, необходимые при работе с тест-системой:
– Пипетки автоматические (диапазон объемов 0,5-1000 мкл)
– Наконечники для автоматических пипеток (до 10 мкл, до 200 мкл, до 1000 мкл)
– Пробирки микроцентрифужные 0,2 мл для ПЦР в реальном времени
– Пробирки микроцентрифужные 0,5 мл
– Пробирки микроцентрифужные 1,5 мл
– Штатив «Рабочее место»
– ПЦР-бокс
– Микроцентрифуга
– Холодильник
– Резиновые перчатки
– Средство для деконтаминации
– Амплификатор ДТ-48 или ДТ-96.
Результаты исследований обработаны методом вариационной статистики по Плохинскому Н.А. (1969), Меркурьевой Е.К. (1970) с использованием ПК и программы «СЕЛЭКС Молочный скот».
2.2 Результаты исследований и их обсуждение
2.2.1 Современное состояние племенной базы крупного рогатого скота костромской породы в племенных хозяйствах Костромской области
Костромская область сложилась как зона молочного животноводства и имеет большой селекционно-производственный опыт. По состоянию на 01.01.2018 г. бонитируемое поголовье Костромской области составило 13582 головы, в том числе, 7165 коров. На долю костромской породы приходится 44,4% всего пробонитированного поголовья области (6034 голов, из которых 3392 головы – коровы).
Племенная база крупного рогатого скота костромской породы представлена тремя племзаводами (ОАО «Племзавод «Караваево» Костромского района, СПК «Гридино» и СПК «Колхоз «Родина» Красносельского района) и двумя племрепродукторами (ООО «Агрофирма «Планета» Буйского района, ООО «Минское» Костромского района).
Сравнительные данные о динамике численности и генеалогическая структура маточного поголовья костромской породы представлены в таблице 7.
Таблица 7 – Генеалогическая структура маточного поголовья костромской породы
| Линия, родственная группа |
Годы | 2018/2014 | ||||
| 2014 | 2015 | 2016 | 2017 | 2018 | ||
| л. Каро КТКС-101 | 18 | 21 | 19 | 17 | 13 | –5 |
| л. Курса ИКС-161 | 116 | 78 | 77 | 62 | 95 | –21 |
| л. Ладка КТКС-253 | 620 | 705 | 826 | 743 | 851 | +231 |
| л. Пика КТКС-419 | – | 4 | 4 | 4 | 3 | +3 |
| л. Салата КТКС-83 | 39 | 24 | 87 | 99 | 141 | +102 |
| р. г. Батлера 107206 | 289 | 176 | 230 | 189 | 137 | –152 |
| р. г. Концентрата 106157 | 496 | 492 | 568 | 654 | 646 | +150 |
| р. г. Мастера 106902 | 598 | 616 | 490 | 598 | 609 | +11 |
| р. г. Меридиана 90827 | 569 | 702 | 691 | 644 | 594 | +25 |
| р. г. Хилла 76059 | 67 | 38 | 45 | 70 | 65 | –2 |
| р. г. Лейрда 71151 | – | – | 43 | 182 | 108 | +108 |
| Прочие линии | 317 | 407 | 200 | 77 | 102 | –215 |
| Общее поголовье | 3129 | 3263 | 3280 | 3339 | 3347 | +218 |
За пять лет, с 2014 по 2018 годы, поголовье скота костромской породы увеличилось на 218 голов (6,9%). Анализируя генеалогическую структуру скота костромской породы, следует отметить, что в целом в породе представлены животные всего 11 линий и родственных групп, непосредственно костромской породы – 5 линий и 6, представляющих родственные группы. При этом такие линии, как Пика КТКС-419 и Каро КТКС-101, представлены единичными животными. Прекратили свое существование линии Силача КТКС-84, Ограды ВДКС-24, Модного КТКС-630, Бархата ВДКС-6, Банана КТКС-333.
В настоящее время наиболее многочисленное маточное поголовье в линии Ладка КТКС-253 (25,4%), родственных групп Концентрата 106157 (19,3%) и Мастера 106902 (18,2%).
Племенные хозяйства области благодаря финансовой поддержке департамента области и федеральным субсидиям стабильно работают и получают неплохие результаты. Производственные показатели коров костромской породы за период 2014—2018 гг. представлены в таблице 8.
Таблица 8 – Производственные показатели коров костромской породы
| Показатели | Годы | ||||
| 2014 | 2015 | 2016 | 2017 | 2018 | |
| Удой, кг | 6300,00 | 6718,00 | 6732,00 | 6664,00 | 6814,00 |
| Содержание жира, % | 4,18 | 4,23 | 4,30 | 4,27 | 4,07 |
| Содержание белка, % | 3,25 | 3,30 | 3,33 | 3,29 | 3,27 |
| Скорость молокоотдачи, кг/мин | 1,62 | 1,51 | 1,73 | 1,88 | 1,94 |
| Возраст коров при 1 отел, дн. | 936 | 941 | 940 | 945 | 955 |
| Сервис период, дн. | 146 | 159 | 151 | 145 | 150 |
| Сухостойный период, дн. | 65 | 68 | 68 | 71 | 70 |
| Возраст 1 случки, мес. | 21,30 | 21,50 | 21,50 | 21,60 | 21,70 |
| Живая масса при 1 осеменении, кг | 403,00 | 416,00 | 409,00 | 405,00 | 420,00 |
| Выход телят на 100 гол. | 85 | 82 | 85 | 84 | 86 |
| Продуктивное долголетие | 3,90 | 4,10 | 4,40 | 3,80 | 3,90 |
За анализируемый период молочная продуктивность коров костромской породы возросла с 6300,00 до 6814,00 кг молока (или 8,2%). Увеличилась скорость молокоотдачи с 1,63 до 1,94 кг/мин.
Возраст коров при первом отеле и срок выращивания ремонтного молодняка увеличились и составили 21,7 месяца и 955 дней против 21,6 месяца и 945 дней в 2014 году. Живая масса телок при первом осеменении возросла с 408,00 кг в 2014 году до 420,00 кг в 2018 году.
Одной из актуальных задач для племенных хозяйств области является сокращение срока выращивания ремонтного молодняка. Показатель по получению телят на 100 голов вырос на 1,0%. В 2018 году существенных изменений по срокам использования коров не выявлено.
В целом приведенные результаты указывают на недостаточные изменения процесса интенсификации и на необходимость активизировать селекционную работу по генетическому совершенствованию скота костромской породы.
2.2.2 Анализ результатов внутрилинейного подбора
Разведение по линиям является основным методом совершенствования пород и стад, так как при этом методе селекционной работы можно осуществить генетически обоснованный подбор.
Линии являются в породе генетической единицей, имеют качественное своеобразие и сохраняют свои наследственные качества в ряде поколений.
Оценка линий костромского скота показала, что лучшие результаты при испытании по потомству имеют производители линий Каро-Курса, Ладка КТКС-253, Мастера 106902, Меридиана 90827, перспективной оказалась и линия Концентрата 106157 (таблица 9).
Таблица 9 – Лучшие результаты подбора в линиях
| Линия, родственная группа | Число
коров гол. |
I лактация | Наивысшая лактация | ||||
| удой,
кг |
жир,
% |
белок,
% |
удой,
кг |
жир,
% |
белок,
% |
||
| Р. гр. Мастера 106902 | 34 | 6117,00± 190,00* |
4,09± 0,03** |
3,27± 0,03 |
7419,00± 204,00 |
4,29± 0,04* |
3,26± 0,03* |
| Л. Ладка
КТКС-253 |
27 | 5508,00± 240,00* |
4,04± 0,04* |
3,39± 0,04 |
6874,00± 302,00* |
4,22± 0,04*** |
3,38± 0,04 |
| Р. гр. Меридиана 90827 | 22 | 6542,00± 380,00* |
3,97± 0,02** |
3,32± 0,02 |
7870,00± 344,00* |
4,19± 0,03*** |
3,30± 0,02* |
| Р. гр. Концентрата
106157 |
33 | 6229,00± 301,00* |
3,81± 0,09** |
3,19± 0,05* |
7151,00± 611,00 |
3,89± 0,04 |
3,17± 0,08* |
| Л. Каро КТКС-101 – Курса ИКС-161 |
20 | 5012,00± 604,00* |
4,05± 0,15 |
3,45± 0,09 |
7654,00± 715,00 |
4,15± 0,14 |
3,50± 0,09* |
| В среднем | 139 | 5633,00± 166,00** |
4,01± 0,03 |
3,30± 0,03 |
7039,00± 132,00** |
4,16± 0,03 |
3,31± 0,03 |
Учитывая то, что чистопородное разведение осуществляется двумя методами спаривания: неродственным (аутбридинг) и родственным (инбридинг), нами проведено исследование происхождения 385 коров, полученных разными вариантами подбора (таблица 10).
Таблица 10 – Молочная продуктивность коров костромской породы при разных вариантах подбора
| Метод подбора |
Число
коров, гол. |
I лактация | Наивысшая лактация | ||||
| удой,
кг |
жир,
% |
белок,
% |
удой,
кг |
жир,
% |
белок,
% |
||
| Аутбридинг | 278 | 5989,00±
87,00 |
4,04± 0,02 |
3,30± 0,02 |
7589,00± 74,00 |
4,19± 0,02* |
3,31± 0,01 |
| Инбридинг | 107 | 6150,00±
125,00 |
4,04± 0,03 |
3,32± 0,03 |
7433,00± 109,00 |
4,27± 0,03* |
3,31± 0,03 |
| В том числе внутрилинейный |
69 | 6102,00±
189,00 |
4,07± 0,06 |
3,35± 0,05 |
7204,00± 203,00 |
4,26± 0,06* |
3,33± 0,06 |
| В том числе: близкий |
3 | 6776,00±
812,00 |
3,83± 0,10 |
3,43± 0,09 |
7892,00± 798,00 |
3,97± 0,11* |
3,27± 0,09 |
| умеренный | 48 | 6307,00±
217,00 |
4,07± 0,04 |
3,37± 0,03 |
7415,00± 235,00 |
4,26± 0,03* |
3,35± 0,02 |
| отдаленный | 56 | 5442,00±
384,00 |
4,10± 0,07 |
3,28± 0,06 |
6530,00± 405,00** |
4,29± 0,06* |
3,24± 0,05 |
Из цифрового материала таблицы 10 следует, что за первую лактацию достоверной разницы по молочной продуктивности между животными, полученными инбредным и аутбредным подбором, нет. Однако отмечаем, что у коров, полученных близким и умеренным инбридингом, их превосходство по массовой доле жира за наивысшую лактацию по сравнению с аутбредными коровами составило 0,07% (Р < 0,05) и 0,10% (Р < 0,05) соответственно.
Низкой продуктивностью отличаются от сверстниц животные, полученные отдаленным инбридингом, на 1059,00 кг (Р < 0,01) молока меньше в сравнении с аутбредными сверстницами.
В племенных хозяйствах использовали в подборе к коровам быков, полученных с применением инбредного подбора: Тика 3818 со степенью инбридинга II-I, Профита 5074 – II-II, Моциона 2162 – III-III и Невода 3033 –IV-III. В таблице 11 приведены данные по молочной продуктивности коров, полученных таким вариантом подбора в сравнении с их матерями, полученными от инбредных быков.
Высокая молочная продуктивность отмечена у дочерей быков, полученных тесным инбридингом. Так, дочери быка Тика 3818 и Профита 5074 имели достоверно выше удои на 1879,00 и 1052,00 кг молока соответственно, чем их матери, что может свидетельствовать о высокой племенной ценности быков, полученных тесным и близким инбридингом.
Таблица 11 – Продуктивность коров, полученных от инбредных быков в сравнении с их матерями
| Кличка и номер быка | Число коров, гол. | Продуктивность матерей | Продуктивность дочерей | ||||
| удой,
кг |
жир,
% |
белок,
% |
удой,
кг |
жир,
% |
белок,
% |
||
| Тик 3818 | 15 | 6255,00±387,00 | 4,20± 0,11* |
– | 8034,00± 278,00* |
4,25± 0,10 |
3,20± 0,01 |
| Профит 5074 | 25 | 7739,00±379,00 | 4,19± 0,12 |
3,47± 0,08 |
8791,00± 242,00*** |
4,23± 0,20 |
3,30± 0,10 |
| Моцион 2162 | 19 | 9579,00±362,00 | 4,13± 0,08* |
3,36± 0,07 |
7667,00± 230,00** |
4,16± 0,07 |
3,22± 0,02 |
| Невод 3033 | 15 | 8630,00±298,00 | 4,23± 0,07* |
3,31± 0,06 |
6992,00± 373,00*** |
4,23± 0,07 |
3,34± 0,04 |
Таким образом, при совершенствовании продуктивности животных за счёт внутрилинейного подбора и подбора сочетанием линий и родственных групп возможно селекционировать животных с высокой молочностью. Однако для этого необходимо усилить внимание к вопросам оценки инбредных животных. Инбредные быки-производители могут эффективно использоваться в племенных стадах для получения коров с высоким уровнем молочной продуктивности.
2.2.3 Сочетаемость линий и родственных групп
При разведении по линиям обращают внимание на их сочетаемость. Сочетаемость линий – это проявление у потомства гетерозиса по продуктивным и хозяйственно-полезным качествам. Сочетаемость линий – генетическая закономерность, обусловленная наследственными особенностями родителей. Поэтому в практике животноводства необходимо учитывать сочетаемость линий, т.е. установить, при скрещивании каких линий получают более продуктивное потомство (таблица 12) .
Высокие показатели молочной продуктивности отмечены у коров, полученных от подбора быков родственной группы Мастера 106902 к коровам, принадлежащим к родственным группам Хилла 76059, Батлера 107206 и линиям Каро КТКС-101 – Курса ИКС-161 и Ладка КТКС-253, как при прямых, так и при обратных кроссах. Так, у коров, полученных при подборе быков линии Каро КТКС-101 – Курса ИКС-161 к коровам родственной группы Мастера 106902 по наивысшей лактации получено 9020 кг молока с содержанием жира 3,96% и 3,40% белка, что на 2559 кг (Р < 0,001) молока больше, чем при сочетании с линией Салата 1216. Дочери быков родственной группы Мастера 106902, полученные от матерей, принадлежащих к родственной группе Концентрата 106157, имели достоверно ниже удой на 913,00 кг молока, чем при обратном кроссе родственной группы Концентрата 106157 и матерей родственной группы Мастера 106902. Таким образом, определены перспективные приемы работы с линиями костромской породы для повышения молочной продуктивности.
Таблица 12 – Сочетаемость линий и родственных групп
| Линия, родственная гр. отца |
Линия, родственная гр. матери |
I лактация | Наивысшая лактация | ||||
| удой, кг | жир, % | белок, % | удой, кг | жир, % | белок, % | ||
| Р. гр. Мастера 106902 | гр. Меридиана 90827 | 5856,00±221,00 | 4,00±0,08 | 3,28±0,06 | 7593,00±271,00 | 4,15±0,09 | 3,28±0,05 |
| гр. Хилла 76059 | 6555,00±361,00 | 4,13±0,05 | 3,39±0,07 | 8133,00±384,00 | 4,46±0,11 | 3,32±0,06 | |
| гр. Батлера 107206 | 6392,00±556,00 | 4,18±0,17 | 3,28±0,01 | 7950,00±477,00 | 4,31±0,18 | 3,65±0,05*** | |
| гр. Концентрата 106157 | 5224,00±432,00 | 4,14±0,15 | 3,12±0,04*** | 6906,00±301,00* | 4,13±0,08 | 3,08±0,07** | |
| л. Каро КТКС-101 | 6576,00±256,00 | 4,16±0,06 | 3,33±0,05 | 8483,00±393,00** | 4,44±0,11* | 3,35±0,05 | |
| л. Ладка КТКС-253 | 6022,00±324,00 | 4,15±0,14 | 3,39±0,11 | 8607,00±399,00** | 4,41±0,15 | 3,31±0,08 | |
| Прочие линии* | 6876,00±410,00* | 3,90±0,13 | 3,25±0,05 | 7838,00±382,00 | 4,08±0,16 | 3,36±0,05 | |
| Р. гр. Меридиана 90827
35 |
гр. Мастера 106902 | 5957,00±198,00 | 4,00±0,06 | 3,38±0,08 | 7468,00±204,00 | 4,20±0,07 | 3,37±0,06 |
| гр. Хилла 76059 | 5856,00±334,00 | 4,20±0,09 | 3,27±0,06 | 7682,00±292,00 | 4,18±0,12 | 3,28±0,05 | |
| гр. Концентрата 106157 | 5854,00±350,00 | 3,98±0,09 | 3,24±0,06 | 6100,00±344,00*** | 4,07±0,10 | 3,27±0,06 | |
| л. Каро КТКС-101 | 5195,00±527,00 | 3,96±0,09 | 3,47±0,04*** | 8418,00±240,00*** | 4,16±0,12 | 3,44±0,07 | |
| Прочие линии* | 652,002±594,00 | 4,08±0,11 | 3,38±0,05 | 8303,00±503,00 | 4,38±0,08* | 3,36±0,07 | |
| Р. гр. Концентрата 106157 | гр. Мастера 106902 | 6649,00±302,00 | 4,05±0,06 | 3,33±0,05 | 7819,00±291,00 | 4,09±0,07 | 3,29±0,05 |
| гр. Меридиана 90827 | 6531,00±377,00 | 4,19±0,07 | 3,35±0,05 | 7376,00±454,00 | 4,32±0,08 | 3,26±0,07 | |
| Прочие линии* | 4699,00±250,00*** | 3,95±0,14 | 3,24±0,04 | 4704,00±248,00*** | 3,98±0,15 | 3,20±0,04 | |
| Р. гр. Лэйрда 71151 | гр. Мастера 106902 | 6404,00±285,00 | 3,99±0,07 | 3,26±0,06 | 7340,00±397,00 | 4,14±0,07 | 3,26±0,05 |
| гр. Меридиана 90827 | 6564,00±758,00 | 3,96±0,13 | 3,27±0,05 | 7349,00±865,00 | 4,23±0,11 | 3,45±0,05* | |
| гр. Хилла 76059 | 6942,00±453,00 | 4,18±0,11 | 3,26±0,04 | 8097,00±671,00 | 4,28±0,15 | 3,33±0,07 | |
| л. Ладка КТКС-253 | 7121,00±545,00 | 4,12±0,14 | 3,27±0,09 | 8777,00±677,00 | 4,14±0,18 | 3,21±0,07 | |
| Р. гр. Хилла 76059 | гр. Мастера 106902 | 5507,00±453,00 | 4,18±0,11 | 3,31±0,07 | 7802,00±450,00 | 4,35±0,11 | 3,29±0,06 |
| Прочие линии* | 4933,00±640,00 | 4,24±0,13 | 3,51±0,09 | 7613,00±710,00 | 4,22±0,12 | 3,34±0,05 | |
| Продолжение таблицы 12 | |||||||
| Линия, родственная гр. отца |
Линия, родственная гр. матери |
I лактация | Наивысшая лактация | ||||
| удой, кг | жир, % | белок, % | удой, кг | жир, % | белок, % | ||
| Р. гр. Батлера 107206 | л. Ладка КТКС-253 | 6672,00±560,00 | 4,02±0,07 | 3,31±0,04 | 8104,00±432,00 | 4,10±0,07 | 3,31±0,03 |
| гр. Меридиана 90827 | 5023,00±647,00 | 3,92±0,11 | 3,20±0,05 | 6661,00±556,00 | 4,12±0,07 | 3,30±0,06 | |
| Прочие линии* | 5025,00±721,00 | 3,95±0,18 | 3,22±0,07 | 5833,00±661,00 | 4,15±0,09 | 3,30±0,11 | |
| Л. Ладка КТКС-253 | гр. Мастера 106902 | 5652,00±388,00 | 4,09±0,12 | 3,35±0,06 | 6693,00±522,00 | 4,22±0,13 | 3,31±0,05 |
| л. Каро КТКС-101 | 5610,00±570,00 | 3,98±0,18 | 3,19±0,05* | 7834,00±575,00 | 4,25±0,11 | 3,33±0,10 | |
| Прочие линии* | 5546,00±548,00 | 3,98±0,15 | 3,36±0,18 | 7649,00±799,00 | 4,34±0,11 | 3,41±0,15 | |
| Л. Каро КТКС-101 – Курса ИКС-161
36 |
гр. Мастера 106902 | 5446,00±538,00 | 3,87±0,15 | 2,82±0,13 | 9020,00±520,00** | 3,96±0,11* | 3,40±0,05 |
| Прочие линии* | 6521,00±489,00 | 3,64±0,18 | – | 6461,00±544,00 | 3,73±0,11 | – | |
| Прочие линии* | Прочие линии* | 5496,00±310,00 | 4,00±0,09 | 3,44±0,08 | 6992,00±482,00 | 4,18±0,10 | 3,60±0,09 |
| В среднем | 6069,00±84,00*** | 4,05±0,02 | 3,31±0,01*** | 7625,00±94,00*** | 4,20±0,02* | 3,31±0,01*** | |
Примечание: * – Прочие линии и родственные группы: линия Салата 1216, быка Тура 2156.
2.2.4 Особенности экстерьера коров костромской породы
При оценке экстерьера коров использовали глазомерный и линейный методы оценки. Комплексная, или классификационная, оценка молочных коров осуществляется по 100-балльной системе (у первотелок максимальный балл – 89).
Данная оценка выявляет животных, наиболее соответствующих желательному типу и используется для индивидуальной оценки коровы и для классификации групп животных с целью их сравнения между собой. Визуальная оценка коров по комплексу признаков применяется для ранжирования животных внутри стада и по породному признаку.
Правильное применение результатов оценки типа телосложения при селекции животных способствует повышению продуктивности коров, легкому протеканию отелов и увеличению продолжительности хозяйственного использования.
В таблице 13 приведены данные оценки коров по комплексу признаков и классификация типа телосложения в баллах с учетом их генеалогической принадлежности. Средний комплексный балл первотелок костромской породы – 77,6 балла, что соответствует классу «хорошо».
Высокий комплексный балл, «хорошо с плюсом», получили первотелки дочери быка Этюда 3696, родственной группы Мастера 106902, Аскета 1760, родственная группа Меридиана 90827, Аврала 1906, родственной группы Концентрата 106157 и швицкого быка-производителя Премьера М 39822332.
При оценке по 100-балльной системе выявлено, что животные родственных групп Меридиана 90827, Концентрата 106157 и дочери швицких быков-производителей имели наиболее ярко выраженные молочные признаки – от 80,50 до 81,40 балла.
Коровы, принадлежащие к родственной группе Концентрата 106157, обладали более глубоким туловищем (80,00) и им же присвоен наибольший балл за конечности – 80,00.
За вымя наибольший комплексный балл присвоен дочерям родственной группы Мастера 106902. Однако следует отметить, что дочери быков-производителей Ключа 872 линии Ладка КТКС-253, Купола 5301 и Наказа 384 родственной группы Меридиана 90827 получили оценку за экстерьер «удовлетворительно».
Им присвоен низкий комплексный балл за конечности и вымя.
Таблица 13 – Оценка экстерьера коров по 100-балльной системе с учетом их генеалогической принадлежности
| Линия/ родственная группа |
Кличка и № быка-производителя | Показатели | |||||
| молочный тип | объем туловища | конечности | вымя | общий балл | оценка | ||
| Л. Ладка КТКС-253 | Ключ 872 | 74,00±0,76 | 77,00±0,54 | 69,00±0,81 | 67,00±0,57 | 70,20 | Уд. |
| Лев 7645 | 75,00±0,57 | 80,00±0,34 | 76,00±0,43 | 80,00±0,68 | 77,70 | Хор. | |
| Марс 289 | 75,00±0,68 | 80,00±0,49 | 79,00±0,37 | 75,00±0,96 | 76,50 | Хор. | |
| среднее: | 76,00±0,13 | 79,20±0,15 | 76,00±011 | 74,20±011 | 75,60±018 | Хор. | |
| Л. Каро КТКС-101
38 Л. Курса ИКС-161 |
Колючий 8810 | 78,00±0,96 | 76,00±1,24 | 75,00±1,15 | 74,00±1,24 | 75,40 | Хор. |
| Сквозняка 6734 | 79,00±0,77 | 80,00±0,82 | 75,00±0,54 | 75,00±0,85 | 76,50 | Хор. | |
| среднее:: | 78,40±0,15 | 77,30±0,12 | 74,90±0,13 | 74,60±0,14 | 75,80±0,16 | Хор. | |
| Р.гр. Мастера 106902 | Лавелас 183 | 81,00±0,45 | 82,00±0,31 | 79,00±0,83 | 77,00±0,87 | 78,00 | Хор. |
| Лед 8556 | 79,00±1,01 | 81,00±0,78 | 78,00±0,87 | 79,00±0,92 | 78,90 | Хор. | |
| Пахарь 157 | 80,00±0,54 | 77,00±0,76 | 81,00±0,98 | 75,00±1,11 | 77,90 | Хор. | |
| Этюд 3696 | 79,00±0,44 | 80,00±0,59 | 81,00±0,67 | 85,00±0,88 | 82,30 | Хор.+ | |
| среднее: | 78,00±0,11 | 78,20±0,10 | 79,00±0,13 | 78,20±0,11 | 78,20±0,16 | Хор. | |
| Р.гр. Меридиана 90827 | Аскет 1760 | 85,00±0,36 | 80,00±0,41 | 80,00±0,74 | 86,00±0,67 | 83,60 | Хор.+ |
| Купол 5301 | 80,00±0,55 | 79,00±0,39 | 71,00±0,84 | 69,00±0,97 | 73,20 | Уд. | |
| Лик 1020 | 81,00±0,65 | 80,00±0,59 | 79,00±0,98 | 75,00±1,04 | 78,00 | Хор. | |
| Лозняк 8093 | 82,00±0,78 | 77,00±0,53 | 78,00±0,81 | 75,00±0,93 | 77,70 | Хор. | |
| Наказ 384 | 79,00±0,21 | 80,00±0,16 | 75,00±0,33 | 69,00±0,39 | 73,10 | Уд. | |
| Среднее: | 81,40±0,14 | 79,20±0,09 | 76,60±0,11 | 74,80±0,09 | 77,30±0,12 | Хор. | |
| Продолжение таблицы 13 | |||||||
| Линия/ родственная группа |
Кличка и № быка-производителя | Показатели | |||||
| молочный тип | объем туловища | конечности | вымя | общий балл | оценка | ||
| Р.гр. Концентрата 106157 | Аврал 1906 | 80,00±0,13 | 79,00±0,11 | 81,00±0,34 | 82,00±0,48 | 80,90 | Хор.+ |
| Леопард 8803 | 82,00±0,67 | 81,00±0,43 | 79,00±0,77 | 71,00±0,84 | 76,70 | Хор. | |
| Среднее: | 81,00±0,08 | 80,00±0,11 | 80,00±0,12 | 76,50±0,15 | 78,80±0,13 | Хор. | |
| Р.гр. Лейрда 71151 | Байкал 8983 | 80,00±0,56 | 81,00±0,43 | 75,00±0,82 | 74,00±0,94 | 76,40 | Хор. |
| Р.гр. Батлера 107206
39 |
Мрамор 6039 | 76,00±0,65 | 75,00±0,71 | 79,00±0,94 | 74,00±0,89 | 75,80 | Хор. |
| Мускат 6061 | 79,00±0,81 | 81,00±0,74 | 78,00±0,89 | 79,00±0,96 | 78,90 | Хор. | |
| Среднее: | 77,50±0,11 | 78,00±0,16 | 78,50±0,10 | 76,50±0,12 | 77,40±0,13 | Хор. | |
| Швицкие быки-производители | Лизол 0206 | 82,00±0,74 | 77,00±0,49 | 78,00±0,95 | 75,00±0,98 | 77,70 | Хор. |
| Мьюзик (Сиде) 198358 | 80,00±1,19 | 82,00±0,98 | 77,00±0,85 | 76,00±0,94 | 77,80 | Хор. | |
| Премьер М 39822332 | 85,00±0,51 | 80,00±0,44 | 80,00±0,34 | 86,00±0,69 | 83,70 | Хор.+ | |
| Среднее: | 80,50±0,11 | 79,70±0,13 | 77,70±0,11 | 76,50±0,14 | 78,10±0,14 | Хор. | |
К дочерям быков-производителей, получивших низкие комплексные баллы, необходимо ставить в подбор быков, дочери которых имеют высокий комплексный балл за конечности и вымя. Это быки Аскет 1760 родственной группы Меридиана 90827, Этюд 3696 родственной группы Мастера 106902, швицкий бык-производитель Премьер М 39822332 родственной группы Концентрата 106157 и т.д. На рисунках 2 и 3 первотелки с высоким комплексным баллом за экстерьер.

Рисунок 2 – Первотелка Пороша 6365, отец бык-производитель Орех 1011,
оценка за экстерьер 82 балла – «хорошо+»

Рисунок 3 – Первотелка Сигара 6382, отец Аврал 1906,
оценка за экстерьер – 83 «хорошо+»
В таблице 14 приведены недостатки экстерьера коров костромской породы в зависимости от генеалогической принадлежности.
Таблица 14 – Недостатки экстерьера у коров костромской породы в зависимости от генеалогической принадлежности
| Недостаток экстерьера | Костромская порода | Швицкие быки-произво-дители
n=7 |
|||||||||
| линия | родственная группа | ||||||||||
| Салата КТКС-83
n=2 |
Ладка КТКС-253
n=6 |
Каро КТКС-101
n=1 |
Курса ИКС-161
n=2 |
Мери-диана 90827
n=7 |
Концентрата 106157
n=4 |
Хилла 76059
n=2 |
Мастера 106902
n=7 |
Батлера 107206
n=3 |
|||
| Мягкая спина | – | 1,18 | 1,18 | – | 1,39 | 4,62 | – | – | – | – | |
| Т
41 |
Горбатая спина | – | – | – | – | – | – | – | – | – | – |
| Слабая поясница | – | 0,59 | 0,59 | – | 0,69 | – | – | – | – | – | |
| Крыловидные лопатки | 6,67 | 37,65 | 37,65 | 6,25 | 37,50 | 23,08 | – | 20,56 | 19,67 | 27,78 | |
| Перехват за лопатками | – | – | – | – | – | 1,54 | – | – | – | – | |
| Итого | 6,67 | 39,42 | 39,42 | 6,25 | 39,58 | 29,24 | 0 | 20,56 | 19,67 | 27,78 | |
| З | Крышевидный зад | – | 2,94 | 2,94 | – | 1,39 | – | – | 2,80 | 1,64 | – |
| Высокий хвост | – | 1,18 | 1,18 | 6,25 | 4,17 | 3,08 | – | 3,74 | 1,64 | 2,78 | |
| Итого | 0 | 4,12 | 4,12 | 6,25 | 5,56 | 3,08 | 0 | 6,54 | 3,28 | 2,78 | |
| Н | Размет передних ног | – | 6,47 | 6,47 | – | 15,28 | 7,69 | – | 17,76 | 8,20 | 2,78 |
| Мягкие бабки | – | 0,59 | 0,59 | – | 1,39 | 1,54 | – | 2,80 | – | – | |
| Большая межкопытная щель | – | 4,12 | 4,12 | – | 7,64 | 6,15 | – | 8,41 | 4,92 | – | |
| Х-обр. | – | 1,76 | 1,76 | 6,25 | 0,69 | – | – | 0,93 | – | 2,78 | |
| Саблистость | 6,67 | – | – | – | – | – | 25,00 | – | – | – | |
| Бочкообр. пост. задних ног | – | – | – | – | – | – | – | – | – | – | |
| Итого | 6,67 | 12,94 | 12,94 | 6,25 | 25,00 | 15,38 | 25,00 | 29,90 | 13,12 | 5,56 | |
| Продолжение таблицы 14 | |||||||||||
| Недостаток экстерьера | Костромская порода | Швицкие быки-произво-дители
n=7 |
|||||||||
| линия | родственная группа | ||||||||||
| Салата КТКС-83
n=2 |
Ладка КТКС-253
n=6 |
Каро КТКС-101
n=1 |
Курса ИКС-161
n=2 |
Мери-диана 90827
n=7 |
Концентрата 106157
n=4 |
Хилла 76059
n=2 |
Мастера 106902
n=7 |
Батлера 107206
n=3 |
|||
| В
42 |
Косое дно вымени | – | 4,12 | 4,12 | 12,50 | 4,17 | 9,23 | – | 3,74 | 3,28 | 5,56 |
| Раздвинутые соски | 6,67 | 1,18 | 1,18 | – | 0,69 | 12,31 | – | 0,93 | – | 11,11 | |
| Сближенные задние соски | – | – | – | – | – | – | – | 2,80 | – | – | |
| Дополнительные соски | 26,67 | 21,18 | 21,18 | 25,00 | 35,42 | 38,46 | 25,00 | 16,82 | 26,23 | 19,44 | |
| Неправ. форма сосков | – | 0,59 | 0,59 | – | 0,69 | 3,08 | – | 1,87 | 3,28 | 2,78 | |
| грушевидная форма сосков | – | 4,12 | 4,12 | – | 2,08 | – | – | 0,93 | – | – | |
| Атрофирование долей вымени | – | – | – | – | – | – | – | – | – | – | |
| Неравномерно развиты доли вымени | – | – | – | – | – | 1,54 | – | – | – | – | |
| Козье вымя | 6,67 | 18,82 | 18,82 | 6,25 | 26,39 | 6,15 | 25,00 | 16,82 | 8,20 | – | |
| Итого | 40,01 | 50,01 | 50,01 | 43,75 | 69,44 | 70,77 | 50,00 | 43,91 | 40,99 | 38,89 | |
При оценке коров костромской породы по экстерьеру были выявлены следующие наиболее часто встречающиеся недостатки – это крыловидные лопатки от 6,25% у коров линии Каро КТКС-101 до 39,58% в родственной группе Меридиана 90827, размет передних конечностей – от 2,78% у швицких быков производителей до 17,76% у коров родственной группы Мастера 106902, козье вымя – от 6,15% в родственной группе Концентрата 106157 до 26,39% в родственной группе Меридиана 90827.
Дополнительные соски с высокой частотой встречаемости от 16,82% до 38,46% выявлены во всех линиях и родственных группах.
Обработка данных по оценке типа телосложения дочерей быков-производителей, используемых в хозяйствах Костромской области, выполнялась в программном обеспечении ОТТ (оценка типа телосложения) ООО «РЦ «ПЛИНОР».
Сведения о быках-производителях и коровах-первотелках по хозяйствам вносятся в программу для дальнейшего включения результатов в каталоги быков-производителей костромской породы.
2.2.5 Основные причины выбытия и продуктивное долголетие коров
С повышением интенсивности отбора в молочных стадах на основе эффективной оценки животных появилась тенденция к получению генетического прогресса за счёт сокращения интервала между поколениями, следовательно, за счёт быстрой смены поколений.
Тем не менее, проблема повышения продолжительности хозяйственного использования молочных коров остаётся актуальной, так как экономическая эффективность отрасли повышается не только с ростом продуктивности, но и с увеличением срока хозяйственного использования коров.
На рисунке 4 приведены основные причины выбытия коров костромской породы.
Наибольшее количество коров выбыло по причине заболеваний конечностей – 23,4%, заболеваний вымени – 18,7%, трудных отёлов –18,7%, послеродовых осложнений и гинекологических заболеваний –16,6%.
По причине внутренних незаразных заболеваний (болезни обмена веществ, заболеваний пищеварительной, дыхательной, кровеносной системы и прочих незаразных болезней, старости, зообрака) выбыло 18,1% , и только 4,5% коров выбыло из стада по причине низкой продуктивности.
Рисунок 4 – Причины выбытия коров
Следует отметить, что в обработку не включались животные, выбывшие из стада по причинам «племпродаж» и «продажа населению». В таблице 15 приведен срок хозяйственного использования коров костромской породы, выбывших по разным причинам.
Таблица 15 – Влияние причин выбытия на продолжительность хозяйственного использования
| Причина выбытия | Число коров | Срок хозяйственного использования, отел | |
| шт. | % | ||
| Низкая продуктивность | 41 | 4,50 | 2,69±0,18 |
| Болезни ног | 212 | 23,30 | 3,35±0,05 |
| Болезни вымени | 170 | 18,80 | 3,87±0,07 |
| Трудные роды | 170 | 18,00 | 3,32±0,07 |
| Болезни половых органов | 151 | 16,60 | 3,25±0,09 |
| Прочие причины | 164 | 18,00 | 3,47±0,14 |
| Итого: | 908 | 100,00 | 3,42±0,03 |
Наибольшее количество коров (23,3%) – выбыло по причине болезни ног, болезни вымени – 18,8% и трудные роды – 18,0%.
Установлено, что четкой закономерности причин выбытия коров из стада в возрастном аспекте не наблюдается.
Так, по причине «трудные роды» средний возраст выбытия – 3,25 отела, по причине болезни конечностей – 3,35 отела и по причине болезни вымени – 3,87 отела. Низкий процент выбытия (4,5%) по причине низкой продуктивности, что свидетельствует о недостаточном количестве ремонтного молодняка в хозяйствах.
Характеристика причин выбытия коров в зависимости от принадлежности к линиям и родственным группам приведена в таблице 16.
Таблица 16 – Причины выбытия дочерей быков-производителей
| Линия/родственная группа | Кличка и номер быка | Число дочерей, гол. | Низкая продук-тивность | Болезни
ног |
Болезни вымени | Трудные роды | Болезни половых органов | Прочие причины | Срок хоз. использ.,
отел |
| Костромские быки-производители | |||||||||
| Л. Ладка
КТКС-253 |
Ключ 872 | 56 | 8,90 | 12,50 | 16,10 | 16,10 | 17,80 | 28,60 | 2,76±0,09 |
| Лев 7645 | 76 | 5,30 | 15,80 | 22,40 | 10,50 | 31,60 | 14,40 | 3,41±0,07 | |
| Л. Каро КТКС-101 – Курса ИКС-161 | Колючий 8810 | 29 | 31,00 | 6,90 | 20,70 | 6,90 | 24,10 | 10,40 | 5,10±0,09 |
| Сквозняк 367 | 15 | 6,70 | 33,30 | 20,00 | 13,30 | 26,70 | – | 4,78±0,06 | |
| Р.гр. Мастера 106902
45 |
Булат 2786 | 45 | – | 20,30 | 11,10 | 13,30 | 11,20 | 44,10 | 4,11±0,13 |
| Лавелас 183 | 47 | – | 21,20 | 17,10 | 19,20 | 19,20 | 23,30 | 4.01±0,07 | |
| Лед 8552 | 19 | 15,80 | 15,80 | 10,50 | 15,80 | 21,00 | 21,10 | 2,71±0,19 | |
| Пахарь 157 | 42 | 4,80 | 14,30 | 14,30 | 11,90 | 21,40 | 33,30 | 4,00±0,09 | |
| Этюд 3696 | 26 | 7,90 | 19,30 | 11,50 | 7,80 | 15,40 | 38,10 | 2,22±0,18 | |
| Р.гр. Меридиана 90827 | Аскет 1760 | 34 | 15,00 | 11,30 | 10,10 | 15,00 | 13,10 | 35,50 | 3,25±0,11 |
| Купол 5301 | 15 | – | 28,50 | 14,30 | 14,30 | 12,40 | 30,50 | 3,10±0,09 | |
| Лик 1020 | 27 | 7,10 | 7,10 | 7,10 | 25,90 | 29,60 | 22,60 | 4,07±0,08 | |
| Лозняк 8093 | 25 | 4,20 | 20,00 | 16,00 | 16,00 | 8,10 | 37,50 | 4,12±0,09 | |
| Наказ 8341 | 46 | 4,30 | 39,10 | 4,30 | 28,30 | 6,50 | 17,50 | 2,90±0,12 | |
| Орех 1011 | 68 | 1,50 | 14,70 | 11,80 | 19,10 | 17,60 | 35,30 | 3,61±0,09 | |
| Р.гр. Концентрата
106157 |
Аврал 1906 | 65 | 3,00 | 21,50 | 10,80 | 35,40 | 6,10 | 23,20 | 3,75±0,07 |
| Леопард 8803 | 54 | 5,50 | 16,70 | 16,70 | 20,40 | 18,50 | 22,20 | 2,56±0,16 | |
| Р.гр. Лейрда 106902 | Байкал 8983 | 27 | 8,30 | 7,60 | 4,30 | – | 5,10 | 66,60 | 6,69±0,19 |
| Р.гр. Батлера 107206 | Мускат 6061 | 39 | 17,90 | 2,60 | 10,30 | 23,00 | 20,50 | 25,70 | 2,50±0,15 |
| Швицкие быки-производители | |||||||||
| Меридиана 90827 | Данкер 173353 | 15 | – | 21,10 | 13,30 | 26,70 | 13,50 | 25,40 | 3,10±0,11 |
| Батлера 107206 | Лизол 0206 | 52 | 1,90 | 21,10 | 17,30 | 17,30 | 17,30 | 25,10 | 4,05±0,08 |
| Концентрата 106157 | Давинчи 9695533 | 51 | – | 19,60 | 3,90 | 11,80 | 13,70 | 51,00 | 2,11±0,15 |
| Эскалибур | 23 | – | 21,70 | 4,30 | 26,10 | 4,30 | 43,60 | 1,74±0,19 | |
| Лейрда 106902 | Перси | 25 | – | 12,00 | 12,00 | 36,00 | 32,00 | 20,00 | 3,00±0,12 |
Как видно из таблицы, самыми непродолжительными были сроки хозяйственного использования дочерей производителей, принадлежащих к родственным группам Батлера 107206, Концентрата 106157, и у дочерей быков родственной швицкой породы: 2,83, 3,18 и 3,25 отела соответственно.
Основная причина выбытия – трудные роды у их дочерей. У быков родственной группы Батлера 107206 выбыли по причинам трудных родов и болезней половых органов 42,5% дочерей. Срок их хозяйственного использования составил 2,23 отела.
У быков-производителей родственной группы Меридиана 90827 и Концентрата 106157 по причине болезней конечностей выбыло 20,2 и 19,4% дочерей соответственно. У швицких быков-производителей выбыли по причинам трудных родов и болезней половых органов 42,0% дочерей, и по причине болезни конечностей – 18,5%.
Выше сроки хозяйственного использования у дочерей быков линии Каро КТКС-101 – Курса ИКС-161, родственной группы Мастера 106902 и Меридиана 908276: 4,97, 3,67 и 3,60 отела соответственно. У дочерей быков костромских линий 23,5% выбыли по причине болезней половых органов и 17,8% – по причине болезней конечностей.
Необходимо проводить ряд профилактически-оздоровительных мероприятий, в первую очередь, пастбищное содержание коров, которое позволяет повысить резистентность организма и продлить срок хозяйственного использования коров. У коров, находящихся на пастбище, значительно реже наблюдаются трудные отелы, задержание последа, парезы и маститы.
Причины выбытия коров по причине болезни вымени зачастую связаны с нарушением технологий машинного доения. Мировой опыт промышленного производства молока показывает, что беспривязное содержание и доение коров в доильных залах имеет преимущество перед доением в стойлах. Поэтому необходимо привязное содержание коров сочетать с автоматической привязью, доением в доильных залах, кормлением на выгульных площадках и выпасом на пастбищах, все это должно соответствовать физиологии животных.
Таким образом, любая современная технология по производству молока должна соответствовать физиологии животного. Своевременная профилактика заболеваний, создание комфортных условий содержания, соблюдение правил доения смогут продлить срок продуктивного использования животных.
2.2.6 Реализация генетического потенциала
На передачу наследственной информации будущему потомству оказывают влияние генотипы не только быков-отцов, но и коров-матерей. В этой связи представляет интерес изучение влияния на качество потомства уровня продуктивности коров-матерей.
Полученные материалы о взаимосвязи удоя, выхода жира и белка в молоке в парах мать – дочь представлены в таблице 17. Установлено большее влияние матерей на выход молочного жира и меньшее – на величину удоя и белка, о чем свидетельствуют представленные в таблице коэффициенты корреляции в парах мать – дочь.
Таблица 17 – Взаимосвязь удоя, выхода жира и белка в молоке коров в парах мать – дочь
| Кличка и номер быка | Кол-во
дочерей в парах мать – дочь |
Коэффициенты корреляции признаков в парах
мать – дочь (r) |
||
| удой,
кг |
жир,
кг |
белок,
кг |
||
| Аврал 1906 | 37 | 0,07 | 0,68 | 0,08 |
| Аскет 1760 | 34 | 0,35 | 0,39 | 0,12 |
| Байкал 8983 | 27 | 0,36 | 0,12 | 0,06 |
| Ключ 872 | 35 | 0,08 | 0,64 | 0,04 |
| Лавелас 183 | 38 | 0,30 | 0,54 | 0,07 |
| Лев 7645 | 28 | 0,15 | 0,49 | 0,11 |
| Лёд 8552 | 21 | 0,27 | 0,67 | 0,11 |
| Леопард 8803 | 17 | 0,29 | 0,24 | 0,08 |
| Марс 289 | 24 | 0,11 | 0,33 | 0,48 |
| Мрамор 3237 | 15 | 0,04 | 0,23 | 0,01 |
| Мускат 6061 | 37 | 0,23 | 0,41 | 0,09 |
| Наказ 3841 | 52 | 0,15 | 0,59 | 0,19 |
| Сквозняк 6734 | 15 | 0,20 | 0,73 | 0,14 |
| Этюд 3696 | 15 | 0,21 | 0,80 | 0,43 |
Низкие коэффициенты корреляции в данном случае подтверждают высокое влияние быков-отцов на основные хозяйственно-полезные признаки коров-дочерей.
Изучение влияния генотипа родителей (матерей и отцов) на величину молочной продуктивности и выхода молочного жира и белка у дочерей за лактацию показало, что сила влияния родительского индекса быка (РИБ) и коровы-матери (РИК) была различной.
На рост удоев коров наибольшее влияние оказали быки-отцы, где сила влияния по РИБ была 0,44, по РИК – 0,19.
В таблице 18 приведены данные продуктивности дочерей быков-производителей костромской породы и разница расчетного показателя РИД с фактической продуктивностью.
Таблица 18 – Продуктивность дочерей быков-производителей костромской и родственной швицкой породы за 305 дней первой лактации и разница расчетного РИД с фактической продуктивностью
| Линия, родственная группа | Кличка быка и номер | Число дочерей, гол. | Продуктивность за 305 дней I лактации |
Реализация генетического потенциала (+/–) |
||||||
| удой,
кг |
жир,
% |
белок,
% |
удой,
кг |
жир,
% |
белок,
% |
|||||
| Быки-производители костромской породы | ||||||||||
| Л. Ладка КТКС-253 | Ключа 872 | 35 | 5261,00±325,00 | 4,09±0,14 | 3,25±0,11 | + 630,00** | + 0,01 | + 0,03 | ||
| Лев 7645 | 28 | 5085,00±319,00 | 3,95±0,14 | 3,26±0,17 | + 562,00* | + 0,23* | – 0,09 | |||
| Л. Каро КТКС-101 – Курса ИКС-161 | Колючий 8810 | 15 | 4557,00±433,00 | 4,36±0,12 | 3,25±0,13 | – 104,00 | + 0,28* | – 0,01 | ||
| Сквозняк 367 | 17 | 4988,00±389,00 | 4,37±0,22 | 3,33±0,12 | + 158,00 | + 0,36* | + 0,11 | |||
| Р.гр. Мастера 106902 | Лавелас 183 | 38 | 5297,00±263,00 | 4,13±0,30 | 3,21±0,09 | + 150,00 | + 0,02 | + 0,08 | ||
| Лед 8552 | 21 | 6238,00±145,00 | 4,28±0,10 | 3,34±0,11 | + 251,00 | + 0,20* | + 0,09 | |||
| Пахарь 157 | 16 | 6011,00±425,00 | 4,21±0,11 | 3,24±0,12 | – 268,00 | – 0,13 | – 0,08 | |||
| Этюд 3696 | 15 | 6084,00±412,00 | 4,35±0,18 | 3,33±0,24 | – 115,00 | + 0,01 | + 0,17 | |||
| Р.гр. Меридиана 90827 | Аскет 1760 | 34 | 6129,00±198,00 | 4,16±0,21 | 3,26±0,14 | + 478,00* | + 0,05 | + 0,04 | ||
| Купол 5301 | 15 | 4777,00±211,00 | 4,01±0,08 | 3.22±0,09 | – 889,00*** | + 0,09 | – 0,13 | |||
| Лик 1020 | 17 | 4790,00±359,00 | 4,49±0,29 | 3,25±0,13 | + 76,00 | + 0,15 | + 0,01 | |||
| Лозняк 8093 | 16 | 5912,00±192,00 | 4,21±0,11 | 3,31±0,12 | + 110,00 | + 0,02 | + 0,01 | |||
| Наказ 8341 | 52 | 4954,00±177,00 | 4,16±0,13 | 3,29±0,09 | – 349,00* | + 0,10 | + 0,08 | |||
| Р.гр. Концентрата 106157 | Аврал 1906 | 37 | 5185,00±144,00 | 4,39±0,29 | 3,29±0,10 | + 109,00 | + 0,16 | + 0,07 | ||
| Леопард 8803 | 17 | 5509,00±217,00 | 4,54±0,11 | 3,37±0,09 | + 630,00** | + 0,22* | + 0,12 | |||
| Р.гр. Лейрда 106902 | Байкал 8983 | 27 | 4950,00±137,00 | 3,94±0,10 | 3,17±0,07 | – 480,00* | – 0,07 | – 0,05 | ||
| Р.гр. Батлера 107206 | Мрамор 6039 | 15 | 5087,00±212,00 | 4,13±0,14 | 3,21±0,09 | – 350,00 | + 0,10 | – 0,04 | ||
| Мускат 6061 | 37 | 5671,00±115,00 | 4,51±0,13 | 3,39±0,12 | + 651,00** | + 0,21* | + 0,06 | |||
| Быки-производители швицкой породы | ||||||||||
| Р.гр. Концентрата | Премьер-М | 29 | 5472,00±233,00 | 4,28±0,22 | 3,29±0,19 | + 1065,00*** | + 0,07 | + 0,13 | ||
| Р.гр. Батлера 107206 | Лизол 0206 | 15 | 5130,00±212,00 | 4,33±0,31 | 3,37±0,24 | + 483,00* | + 0,01 | – 0,02 | ||
| Р.гр. Меридиана | Данкер | 15 | 5915,00±282,00 | 4,38±0,29 | 3,31±0,19 | + 298,00 | + 0,03 | + 0,01 | ||
| В среднем по выборке | – | 511 | 5384,00±31,00 | 4,24±0,01 | 3,31±0,01 | – | – | – | ||
48
Высокая степень реализации генетического потенциала отмечена у костромских быков линии Ладка КТКС-253. Так, дочери быка Ключа 872 имели удой по первой лактации на 650,00 кг (Р < 0,01) молока больше, чем их РИД, а у быка Льва 7545 – на 562,00 кг (Р < 0,05) молока. В родственной группе Меридиана 90827 высокая степень реализации генетического потенциала отмечена у быка Аскет 1760. Его дочери имели удой на 478,00 кг (Р < 0,05) выше, чем РИД. В родственной группе Концентрата 106157 дочери быка Леопарда 8803 превосходили РИД на 651,00 кг (Р < 0,01) молока. В родственной группе Батлера 107206 дочери быка-производителя Муската 6061 имели удои на 651,00 кг (Р < 0,01) молока выше своего расчетного РИД.
Среди швицких быков-производителей высокой степенью реализации отмечен бык-производитель Премьер-М 39822332 родственной группы Концентрата 106157, превосходство над РИД – на 1065,00 кг (Р < 0,001) молока. Более жирномолочными оказались дочери быков Льва 7645 линии Ладка КТКС-253, Колючего 8810 и Сквозняка 367 линии Каро КТКС-101 –Курса ИКС-161 в родственных группах быков Леопарда 8803 и Муската 6061. По показателю белковомолочности достоверной разницы с расчетным РИД не выявлено. Однако следует отметить быка Льва 7645 линии Ладка КТКС-253 превосходство составило 0,09 % по сравнению с расчетным (РИД).
Из вышеизложенного следует, что с целью повышения продуктивных качеств у потомства наиболее предпочтительно использовать быков с высокими показателями продуктивности их матерей и с высокой реализацией генетического потенциала у дочерей.
2.2.7 Мониторинг генетической структуры в породе
Анализ аллелотипа стада должен предшествовать разработке перспективного и текущего планов племенной работы в хозяйстве. Паспортизации по группам крови подлежат: ведущая группа коров и ремонтные телки, бычки, намеченные на племпродажу и для использования в племпредприятиях, дочери быков-производителей, поставленных на оценку по качеству потомства.
Наши исследования показали, что аллелофонд костромской породы претерпевает существенные изменения. Неоспорим тот факт, что использование родственной швицкой породы на животных костромской породы способствовало не только повышению молочной продуктивности скота костромской породы, но и изменению аллелофонда. По сравнению с ранее проведенными исследованиями [88] в породе значительно сократилось количество аллельных вариантов ЕАВ-локуса групп крови, характерных для костромской породы, и повысилась частота встречаемости аллелей, характерных для швицкого скота. В настоящее время отмечается очень высокая степень консолидации наследственного материала, маркированного аллелями G3O1T1Y2E3‘F2‘ и B1O3Y2A’2E3‘G’P’Q’Y’, типичных для швицкого скота американской селекции. Их суммарная частота, встречающаяся в генотипе у коров костромской породы, составляет 0,5209 (таблица 19).
Таблица 19 – Спектр аллелей и частота встречаемости по ЕАВ-локусу групп крови у коров костромской породы
| Аллели ЕАВ-локуса | Частота встречаемости | |||
| 1974—1977 гг. | 2000 г. | 2010 г. | 2018 г. | |
| «b» | 0,2389 | 0,0795 | 0,0846 | 0,0272 |
| Q | 0,1686 | 0,0644 | 0,0231 | 0,0327 |
| О1 | 0,0123 | 0,0076 | 0,0024 | – |
| О’ | 0,1025 | 0,0076 | 0,0513 | 0,0638 |
| I1 | 0,0139 | 0,0516 | 0,0103 | 0,0461 |
| G2D’ | 0,0032 | 0,0303 | 0,0179 | 0,0100 |
| E3‘G» | – | – | 0,0026 | 0,0100 |
| G2О’ | – | – | 0,0096 | 0,0078 |
| А1G’G» | – | 0,0379 | 0,0128 | 0,0100 |
| I1G’G» | 0,1686 | 0,0568 | 0,0410 | 0,0272 |
| I1Y2 Y’ | – | – | 0,0282 | 0,0333 |
| I1Y2E1‘Y’G» | – | – | 0,0154 | 0,0638 |
| Y2I’Q’Y’ | 0,0225 | 0,0114 | 0,0256 | 0,1000 |
| G2E3‘F2‘O’ | – | – | 0,0087 | 0,0039 |
| B1I1T1A1‘P’ | 0,0176 | 0,0076 | 0,0256 | 0,0100 |
| B2P2Y2G’Y’ | – | – | 0,0156 | 0,0100 |
| G3O1T1Y2E3‘F2‘ | – | 0,2195 | 0,3359 | 0,2533 |
| B1G2KE1‘F2‘O’ | 0,0663 | 0,0227 | 0,1051 | 0,0677 |
| B1G2KY2E1‘F2‘O’G» | 0,0573 | 0,0341 | 0,0179 | 0,0093 |
| B1O3Y2A2‘E3‘G’P’Q’Y’ | – | 0,2614 | 0,1359 | 0,2676 |
| Другие аллели | 0,2969 | 0,2705 | 0,0305 | 0,0700 |
| Всего В-аллелей | 42 | 26 | 24 | 20 |
Сложившаяся ситуация с аллелофондом костромской породы в первую очередь объясняется тем, что генетический состав быков-производителей, использовавшихся в стаде, крайне однороден. У 49,0% быков-производителей костромской породы встречается в генотипе аллель G3O1T1Y2E3‘F2‘, три из которых гомозиготны по этому аллелю. Аллель B1O3Y2A’2E3‘G’P’Q’Y’ выявлен в генотипе у 43,0% костромских быков-производителей. Такая высокая частота встречаемости аллелей G3O1T1Y2E3‘F2‘ и B1O3Y2A’2E3‘G’P’Q’Y’ ведет к повышению уровня гомозиготности в стаде и, как следствие, к повышению индекса антигенного сходства при подборе родительских пар.
В последние годы идет тенденция широкого использования быков-производителей, относящихся к родственным группам, и сокращения использования семени быков-производителей костромских линий, что привело к элиминированию и уменьшению частоты ряда аллелей, типичных для скота костромской породы. За период с 1977 по 2018 гг. число аллелей сократилось с 42 до 20. Это обстоятельство вызывает необходимость разработки специальной селекционной программы, обеспечивающей сохранение в стаде необходимого генетического разнообразия с целью сохранения оптимального уровня гетерозиготности, а следовательно, жизнеспособности и продуктивности. Реализация программы по расширению генетического разнообразия аллельных вариантов в стаде позволило бы, по нашему мнению, сохранить многие выдающиеся качества, характерные для костромского скота, и тиражировать их на всю породу.
2.2.8 Полиморфизм генов каппа- и бета-казеинов
Аллельные варианты гена каппа-казеина – одного из основных молочных белков, отличаются нуклеотидными заменами в последовательности цепи ДНК, которые в ряде случаев приводят к изменению аминокислотной последовательности белка, а следовательно, генетически обусловлены.
По данным ученых ВНИИплем, В-аллель гена каппа-казеина указывает на более высокое содержание белка в молоке, а также на лучшие коагуляционных свойств молока коров [37]. По их мнению, наиболее желательным для производства сыров является молоко, полученное от гомозиготных коров с генотипом-ВВ. В таблице 20 приведены данные влияния генотипа по каппа-казеину на молочную продуктивность коров костромской породы.
Таблица 20 – Молочная продуктивность коров с разными генотипами каппа-казеина, X±S
| Генотип | Число коров, гол | Удой,
кг |
Жир | Белок | ||
| % | кг | % | кг | |||
| I лактация | ||||||
| A/A | 14 | 6351,00±321,00 | 4,25±0,08 | 277,00±16,50 | 3,18±0,02* | 201,00±10,20 |
| A/B | 66 | 6364,00±133,00 | 4,15±0,04 | 264,10±5,80 | 3,23±0,01 | 205,00±4,30 |
| B/B | 45 | 6494,00±164,00 | 4,15±0,04 | 269,00±7,10 | 3,25±0,01 | 210,00±5,20 |
| II лактация | ||||||
| A/A | 14 | 6431,00±275,00 | 4,17±0,09 | 268,00±12,90 | 3,21±0,03 | 206,00±8,20 |
| A/B | 61 | 6876,00±125,00 | 4,18±0,04 | 287,00±5,80 | 3,19±0,01 | 219,00±4,10 |
| B/B | 43 | 6789,00±148,00 | 4,18±0,05 | 283,00±6,70 | 3,23±0,02 | 219,00±4,70 |
| III лактация | ||||||
| A/A | 10 | 6676,00±452,00 | 4,15±0,07 | 277,60±19,50 | 3,25±0,04 | 216,30±14,00 |
| A/B | 42 | 7079,00±160,00 | 4,11±0,05 | 291,00±7,70 | 3,20±0,01 | 226,00±5,30 |
| B/B | 30 | 7378,00±202,00 | 4,15±0,06 | 306,00±9,50 | 3,21±0,02 | 236,00±6,10 |
| Наивысшая лактация | ||||||
| A/A | 14 | 7229,00±273,00 | 4,32±0,08 | 312,50±13,10 | 3,22±0,04 | 231,90±7,80 |
| A/B | 66 | 7257,00±123,00 | 4,19±0,03 | 303,00±5,30 | 3,22±0,01 | 233,00±4,00 |
| B/B | 45 | 7302,00±176,00 | 4,15±0,04 | 302,00±7,90 | 3,21±0,01 | 234,00±5,50 |
Из полученных данных видно, что достоверно наибольшим содержанием белка в молоке характеризовались коровы с генотипами AB и BB и только за первую лактацию.
Различия по удою между первотёлками с генотипом AB и AA составили 0,05% (P < 0,05), между первотёлками с генотипом BB и AA – 0,07% (P < 0,01). По другим показателям и по последующим двум лактациям достоверных различий между животными различных генотипов не установлено. Несмотря на отсутствие достоверной разности, всё же удой и содержание молочного белка в среднем за три лактации были выше у коров с теми же генотипами – AB и BB.
По данным племпредприятий АО «Головной центр по воспроизводству сельскохозяйственных животных» за 2018 год и ОАО «Ярославское по племенной работе» за 2018 год проведен анализ быков-производителей по гену каппа-казеина (таблица 21).
Таблица 21 – Генотипы по каппа-казеину быков-производителей племенных предприятий
| Племпредприятие | Порода | Линия/родственная группа | Кличка и номер быка | Генотип по каппа-казеину |
| АО «Головной центр по воспроизводству сельскохозяйственных животных» | костр. | л. Ладка КТКС-253 | Лечо 667 | ВВ |
| костр. | л. Ладка КТКС-253 | Невод 3033 | ВВ | |
| костр. | Л. Каро КТКС-1494 | Сикрон 858 | ВВ | |
| костр. | л. Ладка КТКС-253 | Лексус 695 | АВ | |
| костр. | л. Ладка КТКС-253 | Лакей 463 | АВ | |
| костр. | Р.г. Мастера 106902 | Булат 2786 | АВ | |
| костр. | Р.г. Мастера 106902 | Клык 7834 | АВ | |
| швицкая | Р.г. Лейрда 71151 | Эхо 121 | ВВ | |
| швицкая | Р.г. Лейрда 71151 | Платин 68129 | ВВ | |
| швицкая | Р.г. Лейрда 71151 | Бумер 47579 | АВ | |
| швицкая | Р.г. Лейрда 71151 | Эмпит 79093 | АВ | |
| швицкая | Р.г. Лейрда 71151 | Этап 426 | АВ | |
| швицкая | Р.г. Меридиана 90827 | Индекс 69730 | ВВ | |
| швицкая | Р.г. Меридиана 90827 | Хенк 406 | ВВ | |
| швицкая | Р.г. Меридиана 90827 | Фасон 2655 | АВ | |
| швицкая | Р.г. Меридиана 90827 | Мерлинг 6990 | АВ | |
| швицкая | Р.г. Меридиана 90827 | Дунай 111 | АА | |
| швицкая | Р.г. Меридиана 90827 | Дефис 1 | АА | |
| швицкая | Р.г. Концентрата 106157 | Декрет 85 | ВВ | |
| швицкая | Р.г. Концентрата 106157 | Эскалибур 105117458 | ВВ |
Продолжение таблицы 21
| Племпредприятие | Порода | Линия, родственная группа | Кличка и номер быка | Генотип по каппа-казеину |
| швицкая | Р.г. Концентрата 106157 | Юфаст 47587 | АА | |
| швицкая | Р.г. Ладди | Вексель 61 | ВВ | |
| швицкая | Р.г. Ладди | Взгляд 47 | ВВ | |
| ОАО «Ярославское по племенной работе» | костр. | Р.г. Меридиана 90827 | Залив 9220 | ВВ |
| костр. | Л. Ладка КТКС-253 | Березняк 770 | АВ | |
| костр. | Р.г. Мастера 106902 | Буран 95 | АВ | |
| швицкая | Р.г. Меридиана 90827 | Фармбой 109621953 | АА |
Анализ генотипа быков-производителей костромской и родственной швицкой породы по гену каппа-казеина показал, что из 27 быков 12 (44,4%) имеют желательный генотип ВВ, 11 (40,7%) имеют генотип АВ и четыре быка-производителя (14,9%) с генотипом АА.
Полученные данные свидетельствуют о высокой частоте распространения аллели в популяции скота костромской и родственной швицкой породы. Полученные данные показывают возможность создания референтных популяций с желательным генотипом.
Таким образом, для улучшения технологических свойств молока и повышения продуктивности скота костромской породы желательно использовать семя быков-производителей с генотипом по гену каппа-казеина ВВ и АВ. В целом полученные нами данные согласуются с большинством работ, проведённых по анализу влияния полиморфизма гена CSN3 на показатели молочной продуктивности коров.
В последнее время все чаще исследователи обращают внимание на бета-казеин, роль которого гораздо менее известна зоотехникам. Бета-казеин составляет до 45,0% от общего содержания казеина в молоке. В настоящее время известно более 10 аллельных вариантов бета-казеина, которые можно разделить на группы А1 и А2.
В последнее время наметился устойчивый интерес к так называемому «молоку А2» – молоку, в котором содержится исключительно А2-вариант β-казеина. На волне интереса к молоку А2 многие племенные предприятия стали определять генотип быков-производителей по гену белка β-казеина.
Не исключением стало и АО «Головной центр по воспроизводству сельскохозяйственных животных» (АО ГЦВ) и ОАО «Ярославское по племенной работе», с недавних пор публикующие результаты определения генотипа каждого быка по β-казеину.
В данном исследовании проанализированы данные по 107 быкам молочных и молочно-мясных пород, опубликованные на сайте АО ГЦВ, с методом биометрической статистики. Весь массив протестированных быков были разбит по генам β- и κ-казеина по 9 генотипам (таблица 22).
Таблица 22 – Распределение быков-производителей молочных пород АО ГЦВ по генотипам бета- и κаппа-казеина
| Генотип | Голштинская, ч/п масти
n=76 |
Черно-пестрая
n=12 |
Бурая швицкая
n=11 |
Костромская порода,
n=8 |
|||||
| K-Cas | Бета-казеин | гол. | % | гол. | % | гол. | % | гол. | % |
| АА | А1А1 | 6 | 7,90 | 2 | 16,70 | – | – | – | – |
| АВ | 3 | 3,90 | – | – | – | – | – | – | |
| ВВ | 1 | 1,30 | – | – | 1 | 9,10 | – | – | |
| АА | А1А2 | 12 | 15,80 | 3 | 25,00 | – | – | – | – |
| АВ | 12 | 15,80 | 5 | 41,60 | – | – | 3 | 37,50 | |
| ВВ | 5 | 6,50 | – | – | – | – | 1 | 12,50 | |
| АА | А2А2 | 17 | 22,40 | 2 | 16,70 | 2 | 18,20 | – | – |
| АВ | 13 | 17,20 | – | – | 1 | 9,10 | 2 | 25,00 | |
| ВВ | 7 | 9,20 | – | – | 7 | 63,60 | 2 | 25,00 | |
Как видно из таблицы, на быков с генотипом А2А2 приходится всего 38,4%, при этом наибольшая доля быков с генотипом А2А2 наблюдается в бурой швицкой породе – 90,9%. Другие породы со значительной долей быков А2А2: голштинская – 37,0%, джерсейская – 30,0%, костромская и айширская – по 25,0%. Если же принять во внимание гетерозиготных быков (А1А2), то суммарная доля быков А1А2 и А2А2 выше всего у костромской породы (все 100 % животных).
Несмотря на малое число протестированных быков, следует иметь в виду, что костромскую породу ранее никогда не тестировали по гену CSN-2, и это только первые данные, но они весьма многообещающие. На втором месте по быкам-носителям аллели А2 бурая швицкая порода (90,9%), третье место делят между собой джерсейская и черно-пестрая породы (84,6%), четвертое место у голштинской породы (84,2%).
Если рассматривать распределение массива быков АО ГЦВ одновременно по β- и κ-казеину, то наиболее многочисленная группа – быки с генотипом А1А2АВ – гетерозиготы по обоим генам. На втором месте генотип А2А2АА, на третьем – А1А2АА. Наиболее редкий генотип – А1А1ВВ.
Однако следует признать, что получение стад, полностью состоящих из коров с молоком А2, – это длительная и трудоёмкая зоотехническая работа. В связи с этим необходимо ответить на вопрос: возможно ли отбирать гетеро- и гомозиготных животных по аллели А2, не ухудшая при этом показатели молочной продуктивности?
Для ответа на этот вопрос нами проанализирована молочная продуктивность дочерей быков разных генотипов.
Проведенный анализ влияния генотипа по каппа- и бета-казеину на продуктивность дочерей быков-производителей по сравнению со сверстницами приведен на рисунке 5.
 Рисунок 5 – Оценка племенных качеств быков-производителей разных генотипов
Рисунок 5 – Оценка племенных качеств быков-производителей разных генотипов
На рисунке видно, что только у дочерей быков с генотипами А2А2АВ и А2А2ВВ разность по удою со сверстницами была достоверно выше на 455,00 и 474,00 кг соответственно по отношению с линией среднего значения по всем генотипам.
Таким образом, быки с данными генотипами являются наиболее ценными в селекционном отношении – их использование позволяет получать гомозиготных животных по аллели А2 гена β-казеина с лучшей молочной продуктивностью.
Быки костромской и швицкой породы отобраны для создания референтной популяции с высокими технологическими показателями молока и молока для детского питания (таблица 23).
Дальнейшая работа по созданию референтных популяций с улучшенными качественными показателями молока будет направлена на генотипирование и отбор коров костромской породы с желательными генотипами каппа и бета-казеина.
Создание таких референтных популяций позволит сохранить малочисленные, ценные отечественные породы крупного рогатого скота, неспособные на сегодняшний день конкурировать с обильно молочными импортными породами.
Таблица 23 – Быки-производители, отобранные для создания референтных популяций с заданными параметрами
| Порода | Линия, родственная группа | Кличка и номер быка | Генотип по каппа-казеину | Генотип по
бета-казеину |
| АО «Головной центр по воспроизводству с.-х. животных» | ||||
| костр. | Л. Ладка КТКС-253 | Лечо 667 | ВВ | А2А2 |
| швицкая | Р.гр. Концентрата 106157 | Эскалибур 105117458 | ВВ | А2А2 |
| швицкая | Р.гр. Меридиана 90827 | Хенк 406 | ВВ | А2А2 |
| швицкая | Р.гр. Меридиана 90827 | Индекс 69730 | ВВ | А2А2 |
| швицкая | Р.гр. Лейрда 71151 | Платин 68129 | ВВ | А2А2 |
| швицкая | Р.гр. Лейрда 71151 | Бумер 47579 | АВ | А2А2 |
| швицкая | Р.гр. Концентрата 106157 | Декрет 85 | ВВ | А2А2 |
| швицкая | Р.гр. Ладди | Вексель 61 | ВВ | А2А2 |
| швицкая | Р.гр. Ладди | Взгляд 47 | ВВ | А2А2 |
| ОАО «Ярославское по племенной работе» | ||||
| костр. | Р.гр. Меридиана 90827 | Залив 9220 | ВВ | А2А2 |
| костр. | Р.гр. Мастера 106902 | Буран 95 | АВ | А2А2 |
2.2.9 Генетические аномалии и их контроль
Достижения молекулярной биологии позволяют определять вредные генетические мутации у сельскохозяйственных животных. В отличие от доминантных мутаций, рецессивные остаются в скрытом состоянии в течение многих поколений, не проявляя своего патологического действия. Нами разработана быстрая и чувствительная тест-система для определения рецессивной мутации – дефицит адгезии лейкоцитов (Bovine Leurocyte Adhtsion Deficiency – BLАD).
Принцип действия: определение генотипа основано на проведении полимеразной цепной реакции (ПЦР) с праймерами, общими для «дикого» и «мутантного» вариантов нуклеотидной последовательности:
BLAD-d 5′-TGGCAGGTCAGGCAGTTGCGTT-3′
BLAD-r 5′-CGACTCGGTGATGCCATTGAGG-3′
После завершения температурной программы ПЦР флуоресцентные зонды гибридизуются на полученную матрицу путем понижения температуры реакционной смеси.
Для определения варианта последовательности используется два типа олигонуклеотидов, гибридизующихся на матрицу рядом:
BLAD-pf 5′-CCCCATCGACCTGTACT-(FAM)
BLAD-pv 5′-CCCCATCGGCCTGTACT-(VIC)
BLAD-pq (BHQ)-ACCTGATGGACCTCTCCTACTC-(P).
Первый тип олигонуклеотидов мечен флуорофором, второй – гасителем флуоресценции. Для генотипирования используется один общий олигонуклеотид с гасителем флуоресценции (зонд BLAD-pq) и два сиквенс-специфичных олигонуклеотида, несущих различные флуорофоры: BLAD-pf и BLAD-pv (FAM маркирует предковый аллель, VIC – мутантный аллель). Определение генотипа проводится путем измерения уровня флуоресценции в ходе температурной денатурации дуплексов олигонуклеотидов и полученных матриц. Результаты регистрируются в режиме реального времени.
Если анализируемый образец содержит только один вариант нуклеотидной последовательности гена, т.е. гомозиготен по данному полиморфизму, температура плавления для зонда, образующего совершенный (полностью комплементарный) дуплекс, существенно выше и ниже для зонда, образующего несовершенный (частично некомплементарный) дуплекс (рисунок 6).
Рисунок 6 – Кривая плавления «дикого» варианта нуклеотидной последовательности
При анализе гетерозиготного образца, содержащего оба варианта нуклеотидной последовательности, оба варианта зондов образуют совершенный дуплекс, поэтому температуры их плавления не одинаковы (рисунок 7).
Рисунок 7 – Кривая плавления «мутантного» варианта нуклеотидной последовательности
Разные кривые плавления дают возможность четко разделять образцы на основе их форм.
Специфичность тест-системы оценивали биоинформационным методом, сравнивая последовательность разработанных праймеров против базы данных геномных последовательностей Gen Bank и проверяя их на температурную стабильность и температуру отжига с помощью программного обеспечения Oligo 6.0. По результатам проверки ни один из праймеров не обладал достаточной степенью гомологии с геномом человека, для того чтобы привести к синтезу неспецифической мишени на расчётной температуре отжига.
Чувствительность тест-системы оценивали методом серийных разведений. Чувствительность тест-системы составляет 50 копий ДНК на реакцию. Тест-система демонстрирует стабильную воспроизводимость результатов при анализе количества копий ДНК, превышающего порог чувствительности тест-системы.
Разработанную тест-систему для диагностики дефицита лейкоцитарной адгезии (BLAD) апробировали в племенных стадах Костромской области по разведению костромской, ярославской и черно-пестрой (улучшенной голштинской) породой.
В таблице 24 представлены данные лабораторных исследований.
Таблица 24 – Частота BLAD-мутации разных пород крупного рогатого скота
| Порода | Не носители
TL |
Носители
BL |
Носитель в гомозиготном состоянии | ||
| число | % | число | % | ||
| Костромская | 98 | 100 | – | – | – |
| Ярославская | 96 | 100 | – | – | – |
| Черно-пестрая, «улучшенная» голштинами |
88 | 94 | 6 | 6 | – |
Из данных таблицы следует, что в племенных хозяйствах по разведению отечественных костромской и ярославской пород BLAD-мутации не выявлено. Однако из 94 проб крови черно-пестрой (улучшенной голштинской) породы выявлено шесть процентов животных, носителей этой аномалии.
Проведенная молекулярно-генетическая экспертиза в племенных хозяйствах Костромской области показала, что в черно-пестрой породе, улучшенной голштинами, шесть процентов коров являются носителями аномалии.
Выявление и регистрация животных, являющимися носителями мутации BLAD, будет в большей мере способствовать элиминации этих мутаций из популяции крупного рогатого скота.
Усовершенствованная тест-система BLAD-мутации позволяет своевременно выявлять и регистрировать животных-носителей, что способствует ее элиминации и оздоровлению популяций крупного рогатого скота.
ЗАКЛЮЧЕНИЕ
Получены новые данные по результатам работы с линиями и родственными группами. Определено влияние инбридинга на продуктивность скота костромской породы. Высокая молочная продуктивность отмечена у дочерей быков, полученных тесным инбридингом. Так, дочери быка Тика 3818 и Профита 5074 имели достоверно выше удои на 1879,00 и 1052,00 кг молока соответственно, чем их матери, что может свидетельствовать о высокой племенной ценности быков, полученных тесным и близким инбридингом. Таким образом, тесный и близкий инбридинг рекомендуем использовать только для получения быков-производителей. Остается вопрос о целесообразности применения инбридинга при разведении коров костромской породы.
Анализ результатов сочетаемости линий и родственных групп определил перспективные варианты. Получены новые данные по экстерьеру дочерей быков-производителей и выявлены следующие наиболее часто встречающиеся недостатки экстерьера: крыловидные лопатки от 6,25% у коров линии Каро КТКС-101 до 39,58% в родственной группе Меридиана 90827, размет передних конечностей – от 2,78% у швицких быков производителей до 17,76% у коров родственной группы Мастера 106902, козье вымя — от 6,15% в родственной группе Концентрата 106157 до 26,39% в родственной группе Меридиана 90827.
Дополнительные соски с высокой частотой встречаемости от 16,82 до 38,46% выявлены во всех линиях и родственных группах. Средний комплексный бал первотелок костромской породы – 77,6 балла, что соответствует классу «хорошо». Высокий комплексный балл, «хорошо с плюсом», получили первотелки дочери быка Этюда 3696, родственной группы Мастера 106902, Аскет 1760, родственная группа Меридиана 90827, Аврала 1906, родственной группы Концентрата 106157 и швицкого быка-производителя Премьера-М 39822332. Впервые начата работа по экстерьерному профилированию быков-производителей костромской породы.
Получены данные по основным причинам выбытия. Это заболевания конечностей – 23,4%, заболевания вымени – 18,7%, трудные отёлы – 18,7%, послеродовые осложнения и гинекологические заболевания – 16,6%. Из всего массива коров с высоким продуктивным долголетием 31,0% принадлежит к родственной группе Мастера 106902, 22,0% – к родственной группе Меридиана, по 9,0% – к родственной группе Концентрата 106157 и линии Ладка КТКС-253.
Проведённый генетический мониторинг в стадах костромской породы показал сокращение числа аллелей в два раза. Это обстоятельство вызывает необходимость разработки специальной селекционной программы, обеспечивающей сохранение в стаде необходимого генетического разнообразия с целью сохранения оптимального уровня гетерозиготности, а, следовательно, жизнеспособности и продуктивности.
Анализ влияния белков каппа-казеина на молочную продуктивность скота костромской породы показал, что удой и содержание молочного белка в среднем за три лактации были выше у коров с теми же генотипами – AB и BB.
Анализ генотипа по каппа-казеину быков-производителей костромской и родственной швицкой породы, принадлежащих АО «Головной центр по воспроизводству сельскохозяйственных животных» и ОАО «Ярославское по племенной», показал, что из 27 быков 12 (44,4%) имеют желательный генотип ВВ, 11 (40,7%) имеют генотип АВ и четыре быка-производителя (14,9%) с генотипом АА.
В последнее время все чаще исследователи обращают внимание на бета-казеин, роль которого гораздо менее известна зоотехникам. Установлено, что только у дочерей быков с генотипами А2А2АВ и А2А2ВВ разность по удою со сверстницами была достоверно выше на 455,00 и 474,00 кг соответственно по отношению со средним значением по всем генотипам.
Таким образом, быки с данными генотипами являются наиболее ценными в селекционном отношении – их использование позволяет получать гомозиготных животных по аллели А2 гена β-казеина с диетическими свойствами молока.
В ходе научно-исследовательской работы создана чувствительная и быстрая тест-система для определения мутации BLAD. Система апробирована в племенных стадах Костромской области. В племенных хозяйствах по разведению отечественных костромской и ярославской пород BLAD мутации не выявлено. Однако в пробах крови черно-пестрой (улучшенной голштинской) породы выявлено шесть процентов животных, носителей этой аномалии.
Проведённый анализ позволил выявить и взять на контроль животных с генетическими аномалиями для сокращения животных, носителей данной генетической аномалии.
Акцент на качественные показатели молока, долголетие, устойчивость к ряду заболеваний и поддержка со стороны государства даёт шанс на сохранение и рациональное использование малочисленных отечественных пород.
Список использованных источников
- Абдулина Д.Р., Гизатуллин Р.С., Салихов А.Р. Хозяйственно-биологические особенности коров бурой швицкой породы при чистопородном разведении в условиях Республики Башкортостан // Вестник БГАУ. — 2012. — № 3. — С. 39—40.
- Абрампальский Ф.Н. Оценка типа телосложения и его связь с молочной продуктивностью // Зоотехния. — 2005. — № 10. — С. 2—3.
- Аджибеков К.К., Дунин И.М. Применение метода линейного описания экстерьера животных для оценки коров разных генотипов. Повышение продуктивности отечественных молочных пород путём использования генетического потенциала голштинского скота. — М., 1989. — С. 151—154.
- Айсанов З. Возрастная повторяемость экстерьерных признаков коров // Молочное и мясное скотоводство. — 2004. — № 7. — С. 39.
- Алёшкина C.B. Оптимизация селекции коров на продуктивное долголетие: Автореф. дис. … канд. с.-х. наук. ФГОУ ВПО. — Саранск, 2008. — 25 с.
- Бадин Г.А., Шалугин Б.В. Эффективность выработки твёрдых сычужных сыров из молока коров костромской породы // Актуальные проблемы науки в агропромышленном комплексе: сборник статей 62-й международной научно-практической конференции. — Кострома : Изд. Костромской ГСХА, 2011. — Т. 2. — С. 56—60.
- Бегучев А.П., Безенко Т.И., Боярский Л.Г. и др. Скотоводство. — М. : Агропромиздат, 1992. — 543 с.
- Беляев В.И., Сороковой П.Ф., Машуров А.М. Иммунологический анализ типов крови у коров, устойчивых и предрасположенных к маститу // Генетические методы оценки сельскохозяйственных животных. — СПб., 1985. — С. 47—48.
- Богданов Е.А. Как можно ускорить совершенствование и создание племенных стад (Разведение по линиям). 3-е изд. — М., 1938. — 232 с.
- Богданов Е.А. Что такое породистый скот и как он получается? —М. : Природа, 1918.
- Борисенко Е.Я. Разведение сельскохозяйственных животных. — М. : Колос, 1967. — 463 с.
- Букаров Н.Г. Использование полиморфизма антигенов эритроцитов и главного комплекса тканевой совместимости в разведении и совершенствовании крупного рогатого скота: автореф. дисс. … д-ра биол. наук. (06.02.01). — М. : Дубровицы, 1995. — 33 с.
- Букаров Н.Г., Хрунова А.И., Новиков А.А., Мишина Н.С., Политкин Д.Ю. Оценка быков-производителей по генетическим маркерам групп крови // Зоотехния. — 2010. — № 11. — С. 2—3.
- Бургомистрова О.Н. Взаимосвязь экстерьерных признаков телосложения дочерей быков-производителей с удоем за первую лактацию // Бюлл. ВНИИГРЖ. — СПб., 2006. — Вып. 149. — С. 16—18.
- Валитов Х.З., Карамаев С.В. Иммуногенетические маркеры в селекции крупного рогатого скота по продуктивному долголетию // Известия нижневолжского агроуниверситетского комплекса. — 2011. — № 3 (23). — С. 98—102.
- Всяких А.С. Бурые породы скота. — М. : Колос, 1981. — 271 с.
- Гагиев М.А. Использование экстерьерной оценки типа в селекции бурого скота: Автореф. дис. … канд. с.-х. наук. — ВИЖ : Дубровицы, 1995. — 20 с.
- Гарькавый Ф.Л., Ворслов И.И. Линейная оценка типа бурых латвийских коров // Генетика. — 1991. — № 2.— С. 30.
- Генофонды сельскохозяйственных животных: генетические ресурсы животноводства Росссии / отв. ред. И.А. Захаров; Ин-т общей генетики им. Н.И. Вавилова РАН. — М., 2006. — 426 с.
- Глущенко М.А. Использование генетических методов при оценке заводских семейств костромской породы (06.02.01). Автореф. дисс. … канд. биологических наук. — М. : Лесные Поляны, 1999. — 16 с.
- Гончаренко Г.М., Горячева Т.С., Рудишина Н.М., Медведева Н.С., Акулич Е.Г. Сравнительная оценка сыропригодности молока симментальской и красной степной породы с учётом генотипов k-казеина // Вестник Алтайского государственного аграрного университета. — 2013. — Т. 110.— № 12. — С. 113—117.
- Горский Н.А. Костромская порода скота в колхозах. — М. : Сельхозгиз, 1952. — 172 с.
- Гребень Л.К. Академик М.Ф. Иванов и его работы по выведению новых пород животных. — М. : Учпедгиз, 1949. — 128 с.
- Гринь М.П., Якусевич A.M., Буткевич С.К. и др. Методические указания по линейной оценке типа молочного скота. — Минск, 1998. — 15 с.
- Гумеров У., Исламова С. Аллелофонд групп крови и его связь с продуктивностью коров // Молочное и мясное скотоводство. — 2009.— № 2. — С.11.
- Гусева Т.Ю. Перспективы совершенствования племенной работы с линиями костромской породы // 70 лет костромской породе крупного рогатого скота: сборник статей. — Караваево : Изд. Костромской ГСХА, 2014 . — С. 14—21.
- Донник И.М., Мымрин В.С., Лоретц О.Г. и др. Влияние инбридинга на живую массу коров, экономическая эффективность от инбридинга и рекомендации производству // Аграрный вестник Урала. — 2013. — № 6 (12). — С. 6—8.
- Дунин И.М., Харитонов С.Н., Ермилов А.Н., Сперанский А.Т. и др. Бурый скот России. — М. : Издательство ВНИИплем, 1998. — 101 с.
- Емельянов, Е.Г. Сохранение генофонда крупного рогатого скота костромской породы. — СПб., 2003. — 310 с.
- Ефимова Л. Метод комплексной оценки быков по качеству потомства // Молочное и мясное скотоводство. — 2004. — № 8. — С. 21—22.
- Завертяев Б.П. Тенденции развития методов оценки генотипа животных в молочном животноводстве // Современные методы генетики и селекции в животноводстве // Материалы междун. научн. конф. ВНИИГРЖ от 26—28 июня 2007. — СПб., 2007. — С. 50—54.
- Ивлев Ф.В. Инбридинг и гетерозис в селекции сельскохозяйственных животных. — Кишинев : Картя Молдовеняска, 1987. — 182 с.
- Казаков Д.С. Оценка быков-производителей молочных пород с учётом экстерьера и долголетия дочерей: Автореф. дис. … канд. с.-х. наук. — М. : РАМЖ, 2005. — 30 с.
- Калашникова Л., Ганченкова Т. Оценка холмогорских быков-производителей по генотипу каппа-казеина // Молочное и мясное скотоводство. — 2008. — № 3. — С. 10—12.
- Калашникова Л.А., Дунин И.М., Глазко В.И. и др. ДНК-технологии оценки сельскохозяйственных животных. — Московская область, Лесные Поляны : ВНИИплем, 1999. — 148 с.
- Калашникова Л.А., Дунин И.М., Глазко В.И. Селекция XXI века: использование ДНК-технологий. — Московская область, Лесные Поляны : ВНИИплем, 2000. — 31 с.
- Калашникова Л.А., Труфанов В.Г. Влияние генотипа каппа-казеина на молочную продуктивность и технологические свойства молока коров холмогорской породы // Доклады Российской академии сельскохозяйственных наук. — 2006. — № 4. — С. 43—44.
- Карликов Д.В., Щеглов Е.В., Казарбин Д.Р. и др. Новая система экстерьерной оценки молочного скота // Зоотехния. — 1992. — № 1. — С. 2—5.
- Киселева Н.С. Генетические и физиологические параметры типа «Смоленский» бурого швицкого скота: автореф. дисс. … канд. с-х. наук. —Смоленск, 2005. — 23 с.
- Кисловский Д.А. Избранные сочинения. — М. : Колос, 1965. — 536 с.
- Князева Т.А. Структура популяций красных пород по ЕАВ-локусу групп крови // Зоотехния. — 2016. — № 2.— С.12—13.
- Кольцов Д.Н., Дмитриева В.И. Мониторинг аллелей ЕАВ-локуса групп крови в селекции скота сычевской породы // Зоотехния. — 2011.— № 9. — С. 2—3.
- Кольцов Д.Н., Татуева О.В., Чернушенко В.К. и др. Современное состояние бурой швицкой породы в Российской Федерации и перспективы развития // 70 лет костромской породе крупного рогатого скота: сборник статей. — Караваево : Костромская ГСХА, 2014. — С. 24—32.
- Кравченко Н.А. Племенное дело в животноводстве. — М. : Агропромиздат, 1987. — 285 с.
- Кравченко Н.А. Племенной подбор. — М. : Сельхозгиз, 1957. — 398 с.
- Красильникова Л.H. Методы отбора и подбора при селекции бурых пород скота // Совершенствование бурых пород скота в СССР. — М. : Дубровицы, 1981. — С. 63—70.
- Кугенев П.В., Барабанщиков Н.В. Практикум по молочному делу. — М. : Агропромиздат, 1988. — 224 с.
- Кузнецов В.М. Моделирование селекционного процесса в малочисленных популяциях молочного скота // Аграрная наука Евро-Северо-Востока. — 2005. — № 7. — С. 73—76.
- Кузнецов В.М., Вахонина Н.В. Об ограничении инбридинга в малочисленных популяциях молочного скота // Сельскохозяйственная биология. — 2012. — № 4. — С. 55—57.
- Кулешов П.Н. Выбор по экстерьеру лошадей, скота, овец и свиней. — М. : Сельхозиздат, 1937. — 202 с.
- Кулешов П.Н. Теоретические работы по племенному животноводству. — М. : Сельхозгиз, 1947. — 224 с.
- Лебедько Е.Я. Селекционно-генетические аспекты эволюции бурых пород скота в российском Нечерноземье // Вестник Брянской ГСХА. —2011. — № 1. — С. 3—8.
- Лебедько Е.Я., Непомнящий Г.А. Зависимость продолжительности хозяйственного использования молочных коров от их экстерьерной оценки // Исследования молодых учёных в решении проблемы животноводства. Тез. докл. междун. научн.-практ. конф. — Витебск, 2001. — С. 145—146.
- Легошин Г.П., Агаев Ю.М., Черекаев Н.В. и др. Современная оценка типа телосложения молочных коров // Зоотехния. — 1999. — № 10. — С. 2—6.
- Лискун Е.Ф. Экстерьер сельскохозяйственных животных. — М., 1949. — 312 с.
- Логинов Ж.Г. Роль отбора и подбора в молочном скотоводстве в направлении повышения срока хозяйственного использования коров // Селекционно-генетические методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 2004. — С. 42—49.
- Логинов Ж.Г., Примак В.А., Небасова Н.В. Взаимосвязь полифакторных индексов племенной ценности дочерей с их матерями // Современные методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 2006. — С. 63—70.
- Логинов Ж.Г., Примак В.А., Рахматулина Н.Р. Оценка чёрно-пёстрых коров ленинградского типа по комплексу хозяйственно-полезных признаков // Зоотехния. — 2004. — № 7. — С. 2—5.
- Логинов Ж.Г., Прохоренко П.Н., Дидковский А.Н. Линейная оценка типа потомков голштинских быков при межпородном скрещивании // Современные методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 1991. — С. 61—73.
- Логинов Ж.Г., Прохоренко П.Н., Завертяев Б.П. и др. Улучшение экстерьера молочного скота и повышение срока хозяйственного использования высокопродуктивных коров // Современные методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 2001. — С. 23—29.
- Логинов Ж.Г., Рахматулина Н.Р. Комплексная оценка животных в молочном скотоводстве на основе построения моделей полифакторного индекса племенной ценности // Сельскохозяйственная биология. — 2004. — № 4. — С. 16—23.
- Логинов Ж.Г., Рахматулина Н.Р., Брагинец С.А. Продолжительность хозяйственного использования чёрно-пёстрых голштинизированных коров с различной молочной продуктивностью // Современные методы генетики и селекции в животноводстве. Материалы междун. научн. конф. ВНИИГРЖ. — СПб., 2007. — С. 55—59.
- Логинов Ж.Г., Рахматулина Н.Р., Примак В.А. Комплексная оценка быков-производителей // Современные методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 2001. — С. 55—58.
- Логинов Ж.Г., Семкив М.В. Экстерьерная оценка голштинских быков-лидеров, оставивших наиболее многочисленное потомство в породе // Бюлл. ВНИИГРЖ. — СПб., 1998. — Вып. 145. — С. 18—21.
- Логинов Ж.Г., Шишкина Н.В. Линейная оценка экстерьера голштинских коров // Зоотехния. — 1995. — № 6. — С. 2–5.
- Лось Н.Ф. Влияние типа подбора на молочную продуктивность коров // Зоотехния. — 2003. — №10. — С. 2—5.
- Любимов А.И., Мартынова Е.Н., Азимова Г.В. Оценка молочной продуктивности маточных семейств в ОАО «Племзавод-учхоз «Июльское» Ижевской ГСХА» Удмуртской Республики // Зоотехния. — 2013. — № 8. — С. 2—3.
- Любимов А.И., Юдин В.М. Влияние инбридинга на изменчивость, сопряженность и наследуемость признаков // Достижения науки и техники АПК. — 2013.— № 8. — С. 50—53.
- Лягин Ф.Ф. Эффективность применения инбридинга в селекции высокопродуктивного скота костромской породы. — Караваево : Изд. Костромской ГСХА, 2003. — 46 с.
- Марченко Г.Г. Теоретические вопросы родственного спаривания животных (06.02.01). Автореф. дис. … д-ра с.-х. наук. — М. : Моск. вет. акад. им. К.И. Скрябина, 1992. — 30 с.
- Методические рекомендации по исследованию и использованию групп крови в селекции крупного рогатого скота П.Ф. Сороковой. — М. : Дубровицы, 1974. — 40 с.
- Москаленко Л., Коновалов А. Влияние инбридинга на пожизненную продуктивность коров ярославской породы // Молочное и мясное скотоводство. — 2009. — № 2. — С. 12—13.
- Новиков A.B. Глазомерная оценка статей экстерьера и построение экстерьерного профиля быка по типу его дочерей // Актуальные проблемы интенсификации земледелия и животноводства в современных условиях. Материалы конф. молодых учёных и специалистов. — Свердловск, 1990. — С. 16—20.
- Патент РФ № 2409 945 С1. 21.11.2011. Способ получения коров с высоким сроком продуктивного долголетия // Патент России № 2409 945 С1. 2011. Бюл. № 3 / Еремина М.А., Гриненко А.А., Ахмедова Т.А.
- Попова Н.В. Оценка быков-производителей по экстерьерному типу дочерей // Бюлл. ВНИИГРЖ. — СПб., 1994. — Вып. 141. — С. 6—10.
- Рахматулина Н.Р. Отбор первотёлок по комплексу признаков // Животноводство России. — № 8. — С. 55.
- Рахматулина Н.Р., Егиазарян A.B., Сервах Б.А. Комплексный индекс оценки племенной ценности коров и быков-производителей // Селекционно-генетические методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 2004. — С. 50—61.
- Рахматулина Н.Р., Егиазарян A.B., Сервах Б.А. Оценка генотипа быков-производителей на основе полифакторных индексов // Селекционно-генетические методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. ВНИИГРЖ. — СПб., 2006. — С. 51—57.
- Рахматулина Н.Р., Сервах Б.А., Небасова Н.В. Комплексная оценка генотипа в молочном скотоводстве // Современные методы генетики и селекции в животноводстве. Материалы междун. научн. конф. ВНИИГРЖ СПб. от 26-28 июня 2007 г. — СПб., 2007. — С. 60—63.
- Рахматулина Н.Р., Сервах Б.А., Примак В.А. Динамика изменения показателей экстерьерной оценки коров-первотёлок ГПЗ «Гражданский» // Селекционно-генетические методы повышения продуктивности сельскохозяйственных животных: Сб. научн. тр. / ВНИИГРЖ. — СПб., 2006. — С. 57—62.
- Ржепаковский В.В., Баранник Г.В. Конституция и экстерьер коров чёрно-пёстрой породы и их связь с молочной продуктивностью в учебном хозяйстве института // Сб. научн. тр. — Ставрополь, 1985. — С. 52—54.
- Ружевский, А.Б., Рубан Ю.Д., Бердник П.П. Породы крупного рогатого скота. — М. : Колос, 1980. — 246 с.
- Салбырын Р., Дмитриев В. Анализ способов расчета селекционных индексов // Молочное и мясное скотоводство. — 2008. — № 2. — С. 30—32.
- Сарапкин В.Г., Бялькина Т.А. Комплексная оценка быков- производителей чёрно-пёстрой породы // Молочное и мясное скотоводство. — 2007. — № 5. — С. 4—9.
- Сахаутдинов И., Муратова Л., Исламова С. и др. Аллелофонд групп крови и его связь с молочной продуктивностью симментальских коров // Молочное и мясное скотоводство. — 2011. — № 5.— С. 7—9.
- Свяженина М. Линейная оценка быков-производителей по телосложению дочерей // Молочное и мясное скотоводство. — 2007. — № 4. — С. 21—23.
- Сервах Б.А., Рахматулина Н.Р. Экстерьерная оценка молочного скота // Животноводство России. — 2008.— № 5. — С. 47—48.
- Сороковой П.Ф., Кузьминов И.П. Использование иммуногенетического метода при разведении костромского скота по линиям // Использование иммуногенетических методов в племенном животноводстве: Тез. докл. симпозиума. — Байсогала, 1976. — С. 12—14.
- Стормонт К. Группы крови и их значение в совершенствовании молочного скота // Сельское хозяйство за рубежом. — 1968. — № 6. — С. 24—26.
- Стрекозов Н., Сельцов В., Кожухов Д. Комплексная оценка симменталов поможет селекционерам // Животноводство России. — 2004. — № 11. — С. 16—17.
- Сулимова Г.Е., Лазебная И.В., Перчун А.В. и др. Уникальность костромской породы крупного рогатого скота с позиции молекулярной генетики // Достижение науки и техники АПК. — 2011. — № 9. — С. 52—54.
- Суллер И.Л. Селекция крупного рогатого скота молочных пород : учебное пособие. — СПб. : Проспект Науки, 2012. — 128 с.
- Суровцев В.Н., Галсанова Б.С. Увеличение срока продуктивного использования коров как фактор повышения конкурентоспособности молочного животноводства // Сельскохозяйственные вести. — 2007. — № 1. — С. 11.
- Тамарова Р.В. Создание высокопродуктивных племенных стад ярославской породы крупного рогатого скота // Вестник АПК Верхневолжья. — 2013. — № 1 (21). — С. 31—35.
- Тинаев А., Калашникова Л., Ганченкова Т. Племенные ресурсы быков-производителей чёрно-пёстрой породы // Молочное и мясное скотоводство. — 2009. — № 1. — С. 5—7.
- Ткаченко И.В., Гридин В.Ф. Иммуногенетический маркер жирномолочности коров // Аграрный вестник Урала. — 2014. — № 1 (119). — С. 55—58.
- Тюлькин С., Ахметов Т., Нургалиев М. Технологические свойства молока коров с разными генотипами каппа-казеина // Молочное и мясное скотоводство.— 2011. — № 8. — С. 4—5.
- Тяпугин С.Е. Влияние генетических и паратипических факторов на продуктивное долголетие коров чёрно-пёстрой породы: Автореф. дис. … канд. с.-х. наук. — СПб. : ВНИИГРЖ., 2005. — 25 с.
- Федосенко Е.Г., Баранов А.В., Тараканова Г.Н. и др. Селекция высокопродуктивных коров // Вестник Алтайского государственного аграрного университета. — 2016. — № 2. — С. 78—81.
- Цысь В.И. Повышение эффективности селекции швицкого скота с использованием пород мирового генофонда (06.02.01). Автореф. дис. … д-ра с.-х. наук. Смоленский НИИСХ и СХИ. — М. : Дубровицы, 1995. — 56 с.
- Цысь В.И., Марченко Л.А. Выведение нового молочного типа швицкого скота // Бюл. науч. работ. ВИЖ. — 1991. — Вып. 102. — С. 76—80.
- Цысь В.И., Марченко Л.А. Методы создания высокопродуктивных стад швицкой породы // Повышение продуктивности молочного скота: Сб. науч. тр. — М., 1992. — С. 71—74.
- Цысь В.И., Марченко Л.А. Результаты применения вводного скрещивания при совершенствовании швицкого скота // Матер. научно-практической конференции. — Смоленск, 1994. — С. 109—111.
- Цысь В.И., Марченко Л.А., Чернушенко В.К. Совершенствование швицкой породы с использованием мирового генофонда бурого скота в Смоленской области // Использование пород мирового генофонда при совершенствовании пород отечественного скота: Тез. докл. Всесоюз. научно-практ. конф. — М., 1991. — С. 4—9.
- Цысь В.И., Соколова Е.Г., Леутина Д.В. Качественные показатели молока коров бурой швицкой породы и перспективы их улучшения // Молочная промышленность. — 2015. — № 7. — 16—17.
- Цысь В.И., Стрекозов Н.И., Чернушенко В.К. Российский молочный тип швицкого скота // Ветеринарные и зооинженерные проблемы животноводства: Материалы I международной научно-практической конференции. — Витебск, 1996. — С. 216.
- Чернушенко В.К. Молочное скотоводство Смоленщины в начале XXI века // Зоотехния. — 2009. — № 7. — С. 3—4.
- Чернушенко В.К., Камошенков А., Бабичева В. Тип подбора родителей по ЕАВ-локусу групп крови и хозяйственно-биологические качества дочерей // Молочное и мясное скотоводство. — 2009. — № 2. — С. 9—10.
- Шалугин Б.В. Формирование и реализация продуктивного потенциала скота костромской породы. — Кострома : Изд. Костромской ГСХА, 2011. — С. 199.
- Шаркаева Г. Использование импортного скота на территории Российской Федерации // Молочное и мясное скотоводство. — 2012. — № 1. — С. 12—14.
- Шаумян В.А. Основные вопросы породообразовательного процесса молочного скота. — Кострома, 1948. — 176 с.
- Шишкина Н.В. Оценка племенной ценности быков по комплексу признаков: Автореф. дис. … канд. с.-х. наук. — СПб. : ВНИИГРЖ, 1996. — 20 с.
- Штейман С.И. Как создано рекордное караваевское стадо. — М. : Сельхозгиз, 1948. — 109 с.
- Щепкин М.М. Из наблюдений и дум заводчика // Репринтное издание. Послесловие профессора Е.Я. Лебедько. — Брянск : Издательство Брянской ГСХА, 2006. — С. 42—46.
- Эрнст Л.К., Зиновьева Л.К. Биологические проблемы животноводства в ХХI веке. — М. : РАСХН, 2008. — 508 с.
- Яранцева С.Я. Продолжительность хозяйственного использования черно-пестрых и голштинских черно-пестрых коров разной кровности / диссер. на соискание ученой степени кандидата с.-х. наук (06.02.04). — Новосибирск, 2001. — 116 с.
- Bonfatti V., Cecchinato A., Di Martino G. at all. Effect of к-casein B relative content in bulk milk к-casein on Montasio, Asiago, and Caciotta cheese yield using milk of similar protein composition // J. Dairy Sci. — 2011.— V. 94. — № 2.— P. 602—613.
- Bonfatti V., Di Martino G., Cecchinato A. at all. Effects of beta-kappa-casein (CSN2-CSN3) haplotypes, beta-lactoglobulin (BLG) genotypes and detailed protein composition on coagulation properties of individual milk of Simmental cows // J. Dairy Sci. — 2010. — V. 93. — № 8. — 3809—3817.
- Daetwyler H.D., Daetwyler H., Villanueva B. at all. Inbreeding in genome-wide selection. // J. Anim. Breed. Genet. Bd.— 2007. — 124:369—376.
- Dal Zotto R., De Marchi M., Dalvit C., at all. Heritabilities and genetic correlations of body condition score and calving interval with yield, somatic cell score, and linear type traits in Brown Swiss cattle // J. Dairy Sci. Bd. — 2007. — 90:5737— 5743.
- Fox P., Mullvichil D.M. Milk proteins: molecular, colloid and functional properties // J. Dairy Res. — 1982. — V. 49. — P. 678—693.
- Holt C., Sawyer L. Primary and predicted secondary structures of the caseins in relation to their biological functions // Protein Eng. — 1988. — V. 2. — P. 251—259.
- Ng-Kwai Hang, K.F. Genetic variants of milk proteins and cheese yield // IDF Seminar Cheese yield and factors affecting its control. — Cork, 1993. —P. 160—166.
- Parland S., Kearney J., Rath M. at all. Inbreeding effect on milk production, calving performance, fertility, and conformation in Irish Holstein-Friesians // J. Dairy Science. Bd., 2007. — 90:4411—4419.
- Shuster D.E., Bosworth B.T., Kehrli M.-E.-Jr. Siguence of the bovine CD18-encoding cDNA: comparison with the human and murine glycoproteins // Gene. — 1992. — Vol. 114. — P. 267—271.
- Shuster D.E., Kehrli M.E. Jr., Ackermann M.R. at all. Identification and prevalence of a genetic defect that causes leukocyte adhesion deficiency in holstein cattle // Proc. Nat. Acad. Sci. USA. — 1992. — Vol. 89. — P. 9225—9229.
- Springer T.A. Adhesion receptors of the immune system // Nature. — 1990. — Vol. 346. — P.425—434.
- Tammen I. Weiterentwicklung des DNA-Tests auf BLAD (Bovine Leukozyten Adhasions defizienz) fur den Einsatz in Rinderzucht und klinischer Diagnostik. — Hannover, 1994. — 128 р.
- Thomsen B., Horn P., Panitz F. at all. A missense mutation in the bovine SLC35 A3 gene, encoding a UDP-N- acetylglucosamine transporter, causes complex vertebral malformation // Genome Res.— 2006.— Vol. 16. — P. 97—105.
- Treviranus A. Klinische Befunde und Abstammung von Jungrin-141 dern mit Boviner Leukozyten-Adhäsions-Defizienz (BLAD) / Hannover // Tierarzt. — Hochsch. — 1994. — 152 р.
- Wagner S. Ist Original Braunvich fur Bio besser als Brown Swiss // Bioaktuel. Bd . — 2007. — № 3. — S. 12—13.